Сообщества пустынь
Важными членами экосистем засушливых местообитаний являются термиты (табл. 16.1).
- Термиты
Недавно Johnson и Wood (1979) опубликовали обзор по распространению термитов в аридных зонах Африки и Аравийского полуострова.
Род Psammotermes (четыре вида), относящийся к семейству Phinotermitidae, встречается только в засушливых областях. В Сахаре эти термиты живут в оазисах и по краям пустыни; их гнезда расположены под землей на глубине до 5 м, что позволяет им избегать вредных последствий колебаний температуры, перевевания песка и иссушения. Они могут добывать воду для увлажнения стенок своих гнезд из грунтовых вод на глубине 40 м. Виды Psammotermes фуражируют под прикрытием лепных тоннелей, используя самую разнообразную пищу: навоз, выкопанную почву, подстилку, а в более влажные периоды - древесину.В штате Аризона в горах Санта-Рита на высоте 950 м пять подземных видов потребляют разную пищу при разных способах питания (Haverti, Nutting, 1975; Haverty et al., 1975). Один из них, Heterotermes aureus (Rhinotermitidae), поедает мертвую древесину на поверхности почвы (56% Opuntia, 17% Acacia, 13% Prosopis), тогда как другие термиты предпочитают опунции акацию и поедают также Cercidium или злаки. Lee и Wood (1971) перечисляют 9 подземных видов, обитающих в зарослях кустарниковых эвкалиптов на юге Австралии. В африканской саванне, поросшей туссоковым злаком Loudetia, Bodot (1966) нашел 20 видов термитов, по большей части подземных, хотя некоторые из них строят холмики. Эти виды тоже различаются по своей пище; многие срезают сухие злаки и запасают их (Trinervitermes), другие поедают корни злаков (Amitermes evmcifer). Есть также гумусоядные виды, такие как Cubitermes severus и два вида Trinervitermes, которые кормятся одними и теми же злаками, но используют их по-разному (Bodot, 1966). Bodot описывает также, каким образом тип почвы, влажность и растительный покров влияют на распространение термитов в саванне Берега Слоновой Кости.
В этом районе наблюдается постепенное исчезновение Macrotermes natalensis, возможно в результате вмешательства человека или какого-то изменения климата (или обоих этих факторов). Amitermes evmcifer поселяется в старых разрушающихся холмиках, а также вторгается в холмики Cubitermes severus и некоторых видов Trinervitermes, перестраивая их на свой лад (Bodot,1966). Половые особи, по-видимому, падают на землю и обламывают крылья вблизи этих холмиков, а затем постепенно проникают в них; это относится в особенности к Т. geminatus (Josens, 1972).
Как установил Josens, в Ламто (Берег Слоновой Кости) Macrotermitinae, вероятно, конкурируют за пищу, но не за место для гнезда, поскольку большинство из них строят диффузные подземные гнезда; тем, кто строит холмики, могут помешать найти подходящее место другие термиты; злаки, которыми они питаются, имеются
Таблица 16.1. Обилие и биомасса (живой вес) термитов в различных
системах (Wood, Sands, 1978)
| Экосистема | Вид или группа видов | Тип гнезда1 | Численность на 1 м2 |
Масса,
г/м2 |
Источник данных |
|
Тепло-умеренная зона |
|||||
| Жестколистный лес, юг | Nasutitermes exitiosus | Холмик | 600 | 3,0 | Lea, Wood, 1971b |
|
Австралии |
|||||
| Семиаридный грассленд, | Gnathamitermes | Подземное (1) | 0-9127 | 0-22,21 | Bodine, Ueckert, 1975 |
| Северная Америка | tubiformans | Среднее за 3 года | 2139 | 5,2 | |
| Кустарник / грассленд, Северная Америка |
Heterotermes
aureus |
Подземное (1) | 431 | Haverty, Nutting, Fage, 1975 | |
| Сахельская саванна, Западная Африка | Все виды | Подземное + холмики (2) | 229 | 1,0 | Lepage, 1974b |
|
Тропическая саванна |
|||||
| Саванновое редколесье, север Австралии | Все виды | Подземное (3) | 2000 | Lee, Wood, 1971b | |
| Травяная саванна, Центральная Африка | Cubitermes exiguus | Холмик | 612-701 | 1,3-1,9 |
Hebrant, цит.
no Bouillon, 1970 |
| Саванна | Apicotermes „gurguliflex” | Подземное | 70 | Bouillon, 1962, 1964 | |
| Северогвинейская саванна, Западная Африка |
Trinervitermes
geminatus |
Холмик | 110-2860 | Sands, 1965a,b | |
| Южногвинейская саванна, Западная Африка | Все виды | Подземное + холмик (4) | 4402 | 11,1 |
Wood et al.,
1977 и неопубликованные данные |
| Вторичная южногвинейская | » » | Подземное (5) | 2966 | 3,6 | Wood et al., 1977 |
|
саванна, Западная Африка |
|||||
| Вторичная саванна, Западная Африка | » » | Подземное + холмик (6) | 861 | 1,7 | Josens, 1972 |
|
Тропический лес |
|||||
| Полулистопадный лес, Западная Африка | » » | Подземное (5) | 3163 | 8,0 | Wood, Johnson, неопубликованные данные |
Прибрежный лес,
Центральная Африка Дождевой лес, Вест-Индия Дождевой лес, Южная Америка Дождевой лес, Малайзия
А гроэк осистемы Выпасаемый луг,
Западная Африка Кукуруза (1-й год), Западная Африка Кукуруза (от 8 до 24 лет), Западная Африка
Все виды » »
Nasutitermes costalis Четыре вида
Все виды » »
» »
Пробы почвы брались с глубины 30 см (1), 75 см (2), Josens (1972) и Lepage (1974b) пробы в случае подземных использовались керновые пробы.
Тип гнезда1 Числен- Масса, Источник данных
ность г/м2 на 1 м2
Подземное + 1000
холмик
Подземное (7) 4450
Холмик 87-104
Холмик 1330
Подземное (5) 2010
» » 1553
» » 6825
11,0 Maldague, 1964
Strickland, 1944 0,1 Wiegert, 1970
- Matsumoto, 1976
2,8 Wood et al., 1977
1,7 Wood et al., 1977
18,9 Wood et al., 1977
Рис.
16.5. Горизонтальное распределение грибных садов у двух термитов - Ancistrotermes cavithorax (белые кружки) и Microtermes toumediensis (темные кружки).В центре изображены основания двух деревьев (концентрические кольца). На этом участке находятся две репродуктивные камеры A. cavithorax, возможно, принадлежащие двум разным семьям (большие белые кружки). (Josens, 1972.)
|
•
•1 |
»
0 0 0 щ п ф |
?gt; *%
• V/ # |
f*
Г. amp; |
.... # О
О |
|
•
ф |
•• Ф ¦ * | • # * • • |
%т *
° * * |
т
•• • • * • |
|
0
• •- |
в
$ |
# ¦
і 0 • 8° lt;ъ ’ По Лгgt; |
•
# * |
|
|
о
•lt; • • |
я*
о • о • о |
* s
О |
со ^
0 Ф 0* *- |
90Ф0
• 0 0 • |
|
о
* amp; |
»• |
00
0 * |
%
І •* 000 |
Уlt;
0 |
в изобилии, и 90% обитающих в этой области наземных термитов потребляют злаки. Josens обнаружил здесь 26 видов, из которых 11 встречаются очень редко. Остальные 15 видов можно разделить на следующие экологические группы: наземные-жнецы рода Trinervitermes (четыре вида) и один вид, питающийся древесиной (Amitermes evmcifer); подземные - Ancistrotermes cavithorax, Microtermes toumediensis, Odontotermes (вид неизвестен) и Pseudacanthotermes militaris, выращивающие грибы, один вид, питающийся древесиной без участия грибов (Microcerotermes parvulus), и пять гумусоядных видов (Josens, 1972). Идентифицировать отдельные семьи Macrotermitinae не удалось-имеются лишь косвенные сведения о существовании семей, и возможно, что они слились; однако Josens произвел подсчет среднего числа грибных камер на гектар и получил следующие данные: Ancistrotermes cavithorax -21 500, Microtermes toumediensis - 24 400, Odontotermes-3300 и Pseudacanthotermes militaris -8000.
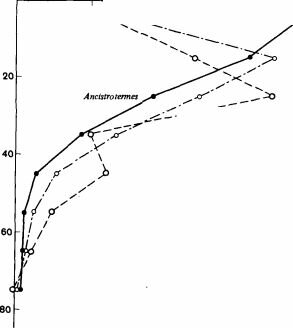
Были получены также подробные сведения о составе этих популяций, их биомассе и превращении энергии (рис. 16.5 и 16.6). После ежегодных пожаров растительных остатков бывает мало, хотя кустарники и деревья сбрасывают листву; в таком случае между термитами должна быть конкуренция, несмотря на их диффузные гнезда, распределение по разным ярусам и различия в питании. Из 9920 приманок (связок травяных стеблей), расположенных в шахматном порядке, только 40 использовались одновременноРис. 16.6. Вертикальное распределение грибных камер (в процентах от их общего числа) четырех видов термитов: Ancistrotermes cavithorax; Odontotermes spp.; Pseudocanthotermes militaris и Microtermes toumediensis вместе. (Josens, 1972.)
двумя видами (Ancistrotermes cavithorax и Microtermes toumediensis). Поскольку в одиночку A. cavithorax использовал 2485, a M. toumediensis -2059 приманок, то, если эти виды безразличны друг другу, число общих приманок должно было бы составить 511. В тех случаях, когда оба вида используют одну и ту же приманку, они устраивают земляную перегородку, чтобы отделиться друг от друга.
Хотя примеры сообществ термитов и их описания многочисленны (см. Krishna, Weesner, 1970), прямых данных о взаимодействиях термитов мало. Согласно Noirot (1959), по крайней мере разные роды избегают встреч, a Bouillon (1970) приводит интересный пример взаимодействия, когда наличие гнездового холмика Macrotermes
10 20 30
—I
Pseudacan th о term es О. о-— и Microtermes
Грибные сады на каждой глубине, %
Odontotermes
natalensis явно препятствует устройству гнезда Cubitermes sankurensis в радиусе 5-10 м. Такого рода конкуренция не обязательно приводит к схваткам; она связана с тем, что пища используется более эффективно, если ее источник находится вблизи гнезда. Hodotermes mossambicus и Trinervitermes trinervoides питаются злаками в одной и той же местности и обычно избегают друг друга, а если их насильственно смешать, то между ними возникают драки (Nel, 1968).
В южной Австралии разные виды Coptotermes обитают в разных бревнах (Calaby, Gay, 1956, цит. по Lee, Wood, 1971). Плотность Nasutitermes exitiosus достигает 8,6-9,2 холмиков на гектар, а при наличии в данной местности Coptotermes lacteus-всего 2,4 на гектар; они рассеяны по занимаемой ими территории. У трех родов термитов, питающихся злаками, тоже наблюдается тенденция к дисперсии. Nasutitermes triodei в присутствии Hemitermes laurensis чаще покидает свои холмики; оба эти вида питаются злаками, и хотя первый из них поедает, кроме того, помет животных и растительные остатки, это его, очевидно, не спасает (Lee, Wood, 1971).Сходные сообщества частично наземных термитов изучал Josens (1972) в Ламто. Он нашел на 1 гектаре в среднем 40,5 гнезда Trinervitermes geminatus (60% гнезд
и 80% всей биомассы термитов), 14,7 гнезда Т. togoensis, 3,2 гнезда Amitermes evuncifer, 2,6 гнезда Т. oeconomus и в меньшем числе гнезда двух других видов Trinervitermes. В травяной саванне (главным образом Loudetia) есть бугорки, образовавшиеся из старых распадающихся холмиков Macrotermes; на этих хорошо дренируемых островках обосновывается Т. togoensis, получая таким образом возможность селиться в сырых, покрытых травой местах.
Т. geminatus обитает на более открытых, но поросших кустарником участках, а численность Т. oeconomus достигает максимума в очень густой кустарниковой саванне. Гнезда Macrotermitinae, ведущих подземный образ жизни, настолько диффузны, что, по мнению Josens, они конкурируют только за пищу; что же касается наземных видов, среди которых 9/10 составляют термиты-жнецы, то, как показывают его расчеты, из 4-5,5 т/га (по сухому весу) общего количества собранного ими корма съедается только несколько десятков килограммов.
Sand (1965), изучавший Trinervitermes в саваннах Нигерии, уделял главное внимание сравнению видов, принадлежащих к одному роду, а не исследованию сообществ; тем не менее интересно отметить, что, как он обнаружил, в гнездах Trinervitermes обычно живут и другие термиты или же муравьи, а иногда те и другие. Обычно никаких контактов между разными видами как будто бы не бывает, но если вскрыть холмик, то „немедленно возникает сумбурный конфликт".
В гнездах Trinervitermes trinervius живет один вид Microtermes, в гнездах Macrotermes natalensis- другой вид, но хозяева не обращают на них внимания. Ввиду резкого различия в размерах между сожителями эти взаимоотношения можно, по-видимому, считать типичным случаем парабиоза.
La Fage и Nutting (1978), изучая взаимоотношения термитов в пустыне вблизи Таксона (Аризона), помещали бумажные приманки либо на сеточки, либо под поверхностный слой почвы. Термиты Heterotermes aureus прокладывали к 53% этих приманок узкие неровные галереи, a Gnathotermes perplexus - более широкие и аккуратные галереи к 96% приманок; к тем приманкам, которые использовали оба вида, каждый из них подходил по своим галереям, избегая галерей, проложенных другим видом.
Термиты нередко поселяются в гнездах других видов в качестве сожителей. Вид-сожитель часто бывает гораздо мельче вида-хозяина и менее активен; такие два вида могут быть либо родственными, либо очень далекими (Bouillon, 1970). Иногда этот парабиоз бывает облигатным, оба вида могут пользоваться одними и теми же галереями, и их репродуктивные пары находятся в тесной близости, а в других случаях вид-хозяин изолирует от сожителя свою царскую пару. Josens
- обнаружил вокруг камеры термита Ancistrotermes cavithorax и в ее стенках галереи значительно более мелкого гумусоядного термита, а также аналогичные галереи в стенках репродуктивной камеры Pseudacanthotermes militaris. Помимо такиях явных примеров использования одним видом гнезда другого вида термиты, строящие большие холмики, разрыхляют почву, помогая другим видам проникать в нее. В Южной Америке Comitermes cumulans строит в грассленде холмики высотой до 4 м. Если эти термиты покидают свои холмики, то в них поселяются общественные осы, муравьи и даже рептилии, а также другие термиты. Sands в своих экспериментах показал, что некоторые виды термитов предпочитают землю, взятую из холмика другого вида, обычной почве. Термиты размельчают не только почву; они могут размельчать древесину, чтобы снабжать гумусоядных термитов перегноем, создавая тем самым сукцессию от видов, питающихся древесиной, к гумусоядным видам.
На термитов часто охотятся муравьи; Camponotus, Crematogaster и Pheidole поедают Trinervitermes. Megaponera foetens не только охотится на многих Macrotermitinae, но и поселяется в их старых холмиках. В районе Моквы (9° с. ш.) из 83 изученных гнезд этого муравья 37 находились в заброшенных холмиках Macrotermes bellicosus, 3-в холмиках Trinervitermes, 36-в земле под кустами и 7-на открытых местах (Longhurst, Howse 1979а, b). Доля общих гнезд с Trinervitermes geminatus варьирует в зависимости от местообитания и возрастает там, где много старых холмиков. При парабиозе муравьи обычно избегают термитов и безусловно не нападают на них, пока термитник не поврежден. Многие Dorylinae истребляют термитов, солдаты которых либо оказывают им сопротивление, защищая шаг за шагом свое гнездо от проникновения муравьев, либо уходят колоннами, двигаясь по поверхности земли, и возвращаются после прекращения атаки. Апотта kohli может затратить несколько дней, чтобы проникнуть в большой холмик Cubitermes. Среди термитов Macrotermes bellicosus в Мокве, где их многочисленные холмики беспорядочно разбросаны по саванне, очень высока смертность от нападения хищников, и, как считает Collins (1981а), их плотность регулируется скорее хищниками, чем внутривидовой конкуренцией.
На одном гектаре в Ламто (Берег Слоновой Кости) можно обнаружить
- млн. собирающих или срезающих траву термитов вместе с 4,5 млн. особей гумусоядных видов и 5 млн. термитов, выращивающих грибы (Josens, 1972). Фуражиры или жнецы потребляют 30—50 кг сухой травы в год, гумусоядные- примерно 30 кг целлюлозы в год и перекапывают не менее 15 тонн почвы за год, тогда как виды, выращивающие грибы, ежегодно вносят в почву около 1,3 тонны растительных остатков (сухой вес).
Еще по теме Сообщества пустынь:
- Продуктивность сообществ и жизненные стратегии водорослей. Морфофункциональный подход к изучению сообществ макрофитов
- СОСТАВ СООБЩЕСТВ И КОМПЛЕКСЫ СООБЩЕСТВ
- РАСТИТЕЛЬНОСТЬ ПУСТЫНЬ И ПОЛУПУСТЫНЬ
- Пустыни умеренной зоны
- Муравьи пустынь
- Серая пустынная почва, серозем.
- «КОРАБЛИ ПУСТЫНИ»
- РАКИ В ПУСТЫНЕ
- Рифовые сообщества Особенности рифовых сообществ
- Гетеротрофные сообщества
- Ассоциация и сообщество
- Изучение структуры сообществ
- Параллельные сообщества. Трофические зоны
- ФУНКЦИОНАЛЬНОЕ ОПИСАНИЕ СООБЩЕСТВ
- СПЕЦИФИКА ПЕЛАГИЧЕСКИХ СООБЩЕСТВ
- ОРГАНИЗАЦИЯ И ТИПЫ СВЯЗЕЙ В БИОТИЧЕСКОМ СООБЩЕСТВЕ
- РАЗДЕЛЕНИЕ ЖИВОГО ПОКРОВА ЗЕМЛИ НА СООБЩЕСТВА [23]
- 13.1.3 Свет в растительном сообществе
- Практика и традиции выделения сообществ макробентоса
- Гидротермы и другие сообщества хемобиоса