Рифовые сообщества Особенности рифовых сообществ
Слово «рифы» обычно ассоциируется со словом «коралловый», хотя не все рифы — коралловые и не все кораллы образуют рифы. Коралловые рифы — одни из наиболее богатых (как в смысле числа видов, так и создаваемой продукции) сообществ Земли.
Основу богатства коралловых рифов обеспечивают кораллы, имеющие симбиотические водоросли. Именно поэтому сообщества коралловых рифов рассмотрены в разделе автотрофних сообществ. С другой стороны, коралловые рифы можно рассматривать как сообщества фильтраторов эпифауны. В некоторых из них доминируют автотрофные виды, в других нет, но в их функционировании и эволюции много общих черт, поскольку рифовые сообщества эволюционно возникают из сообществ фильтраторов эпифауны. Это делает логичным рассмотрение в этом разделе всех рифовых сообществ, а не только тех, в которых высока доля фотосинтетиков.
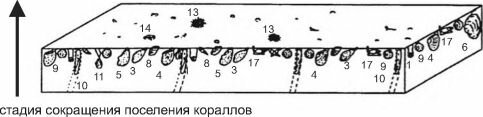
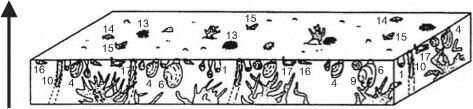
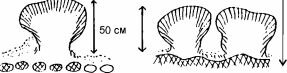
Все виды, способные к доминированию в сообществах фильтраторов эпифауны, во взрослом состоянии мало- или вовсе неподвижны. Все фильтраторы в процессе своей жизнедеятельности продуцируют фекалии и псевдофекалии. В результате на дне накапливается ил и детрит. Накоплению мелких фракций также способствует снижение скорости движения воды в сообществах фильтраторов, особенно у грунта, подобно тому как скорость ветра падает в лесу. Это приводит к ухудшению условий существования фильтраторов, в пределе — к заносу их илом и смену сообщества фильтраторов эпифауны сообществом фильтраторов инфауны. Пример такого развития событий показан на рис. 3.16: постепенно накапливающийся ил погребает кораллы, некоторые вершины их колоний ещё возвышаются над осадком на стадии сокращения поселений кораллов, на последней стадии накапливающийся осадок погребает и их.
Самые простые сообщества фильтраторов эпифауны — мидиевые банки. Мидии способны образовывать практически одновидовые сообщества со 100% покрытием. В этих условиях оседание новых мидий возможно только на других мидий, но этому препятствует фильтрующая деятельность мидий, отфильтровывающих своих личинок и переводящих их в псевдофекалии.
В результате мидиевые банки стареют. Поскольку мидии при-
![]()
![]()
крепляются к субстрату с помощью биссуса, после гибели моллюска створки отваливаются от субстрата. Поэтому мидии не способны к строительству трёхмерных рифов и в местах их доминирования со временем образуется заиленная ракуша, на которую прикрепляются моллюски. Примером такого сообщества является биоценоз мидиевого и фазеоли- нового илов Чёрного моря, но, обычно, мидиевые банки сменяют сообщества другого состава (Ошурков, 2000). Устрицы прирастают к субстрату нижней створкой, но верхняя, на которую прикрепляются другие фильтраторы, после смерти устрицы отваливается. Поэтому устричные банки способны только к медленному росту в высоту и фактически не превращаются в рифы.
Единственный способ уменьшить негативный эффект заиления — подъём фильтраторов над осаждающимися илом и детритом, т. е. над поверхностью грунта. Проще всего это достигается увеличением индивидуальных размеров (особей или колоний) и развитием приспособлений для подъёма организма над поверхностью грунта. По этому пути пошли фильтраторы эпифауны рыхлых грунтов, развитие сообществ которых началось ещё в кембрии. Однако такая жизненная форма чувствительна к подвижности осадков и труднозащитима от хищников. Подвижность осадков прямо пропорциональна интенсивности гидродинамики, что при прочих равных условиях равноценно скорости приноса пищи водой. Т. е. в наиболее кормных условиях фильтраторы эпифауны рыхлых грунтов оказываются в наихудших условиях в отношении размещения их на грунте.
Образование трёхмерных структур снижает чувствительность сообществ фильтраторов эпифауны к заилению — непременному побочному эффекту фильтрации. Поэтому неудивительно, что такие сообщества появились уже в раннем кембрии. Для нерифовых сообществ в англоязычной литературе есть даже специальный термин — level-bottom community — сообщества не возвышающиеся над поверхностью грунта.
Строители настоящих рифов — это или колониальные организмы, или организмы, образующие скопления. Колониальными являются многие кишечнополостные, губки, мшанки и вымершие строматопороидеи. Рифовые скопления способны образовывать известковые водоросли (см. выше), губки, брюхоногие моллюски (Vermetidae, Turritellidae (род Vermicularia) и Siliquariidae), полихеты с известковыми (Serpulidae) или песчаными
![]()
30 см
1
2
![]()
5 см ![]()
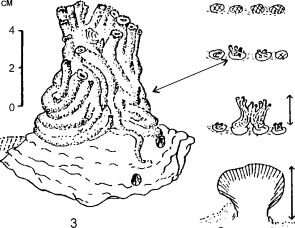
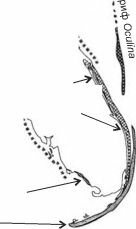
![Рост сабелляриевых рифов на разных субстратах и строение отдельных колоний]()
Рис. 3.54. Рост сабелляриевых рифов на разных субстратах и строение отдельных колоний. По
Les recifs... (1988).
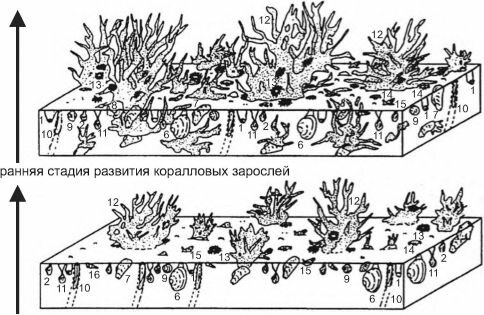
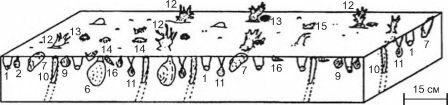
стадия максимального развития коралловых зарослей
![]()
время
начало колонизации кораллами двустворчатых моллюсков
![Изменение состава сообществ при развитии поселений ветвистых кораллов на мелководье у атлантических берегов юга Северной Америки в плиоценовое время]()
Рис. 3.53. Изменение состава сообществ при развитии поселений ветвистых кораллов на мелководье у атлантических берегов юга Северной Америки в плиоценовое время. По Bailey, Tedesco (1986) из JI.A. Невесской (1999: 330).
Таксоны: 1 — Parvilucina, 2 — Carvocorbula, 3 — Macrocallista, 4 — Spisula, 5 — C.rassatella, 6 — Mercenaria, 7 — Modiolus, 8 — Noetia, 9 — C.yclocardia, 10 — Ertsis, 11 — Cumingia, 12 — Septastrea, 13 — Arbacia, 14 — Panopeus, 15 —Nassarius, 16 — Turritella, 17 — Olivella.
риф Sabellaria vulgaris
w?.
![]()
риф
Sabellaria floridens
Florida Middle ground
риф Vermetidae
![Распределение рифов у берегов Флориды.<div class=]()
По Репу (1992)" />
*• f) риф.
• j Phragmatopomma^ \r"j caudata
тропический коралловый риф
Рис. 3.55. Распределение рифов у берегов Флориды. По Репу (1992).
(Sabellariidae: Sabellcirici, Phrag- matopoma) (рис. 3.54) трубками.
Они создают прочный каркас (обычно известковый), скорость формирования которого превышает скорость осадконакопле- ния. Эта способность и делает возможным образование рифов.
Рифы, образованные разными организмами, распределяются закономерно. На рис. 3.55 показано распределение рифов у берегов Флориды. Риф Oculina образован Oculina varicosa и располагается на глубине 70-100 м; этот вид кораллов — один из немногих, способных нормально жить как с эндосимбионтами, так и без них (Curtis, Stanley, 1981). Рифы Sabellariidae образованы вдоль восточного побережья Флориды Sabellaria vulgaris севернее и Phragmatopomma caudata южнее, а вдоль западного побережья — S.floridens. Длина рифа Ph. caudata — около 300 км, высота до 1 м. Vermetidae, образовавшие рифы у юго-западных берегов Флориды, по-видимому вымерли в данном месте в конце XIX века, но образованный ими риф до сих пор вполне отчётлив (Репу, 1992).
Обычно, говоря о коралловых рифах, подразумевают коралловые рифы тропических мелководий. Коралловый риф в наиболее привычном понимании представляет собой противостоящую прибою известковую конструкцию на мелководных платформах тропических морей. Толщина кораллового известняка по результатам бурения может достигать 2 км и более.
Коралловые рифы представляют собой одно из наиболее сложных и продуктивных сообществ не только Мирового океана, но и Земли в целом. При этом коралловые рифы существуют в казалось бы бесплодных местах. Мелководные коралловые рифы омывают воды с содержанием биогенов в следовых количествах: содержание Р04-Р в среднем составляет 0,1-0,4 мгк ат./л, что примерно соответствует уровню лимитирования роста для фитопланктона (0,2 мкг ат./л), а азот в значительной степени фиксируется в пределах рифа.
Коралловые рифы расположены в водах с различным уровнем продуктивности и содержания биогенов. Однако концентрация биогенов внутри мелководных рифов мало зависит от из концентрации вне рифа (Smith, 1985, цит. по: Сорокин, 1990). Это свидетельствует о том, что экосистема рифа сама регулирует концентрацию биогенов в воде внутри рифа. В значительной степени биогены сообщество коралловых рифов получает из растворённого органического вещества, причём из наиболее стойких органических веществ: гуминовых кислот и лигнинов — тех, которые придают коричневую окраску воде наших болот. Имеются данные, что эти вещества разрушаются только на коралловых рифах, т. е. эти биогены недоступны для прочих сообществ. Происходит это окисление на поверхностях раздела фаз (пористый известковый субстрат и песок) (Сорокин, 1990). Способность усваивать биогены из недоступных другим сообществам источников и эффективно их удерживать внутри сообщества приводит к тому, что биологическая продукция коралловых рифов на 2 порядка выше, чем в окружающей пелагиали. Как минимум на порядок активнее и бактериальная микрофлора. Общую площадь мелководных коралловых рифов
оценивают в 0,2% от площади Мирового океана (Kleypas, 1997), из которых 30% к настоящему времени уже сильно повреждены, ещё 30% находятся в угрожающем состоянии.
Это тем более печально, что коралловые рифы — одни из основных сообществ Земли, связывающие углекислый газ: скорость роста рифа до 150 мм/год, в год все мелководные рифы связывают 2,5 млрд. т карбонатов (Сорокин, 1990), что в пересчёте на углекислый газ дает 1,1 млрд. т. Именно их с полным основанием можно назвать «лёгкими планеты», а не леса, как это часто утверждают в популярной литературе. В самом деле, функция лёгких — газообмен, т. е. удаление углекислого газа и поставка кислорода. Эти процессы связаны простым балансовым уравнением фотосинтеза, из которого явствует, что одновременно с выделением кислорода должен связываться углерод, т. е. происходить явственное накопление веществ, содержащих углерод.
В лесах весь поглощённый углекислый газ сохраняется в виде тел организмов, да в виде незначительного количества гумуса. Поскольку накопления связанного углерода ни в какой форме (биомасса, гумуса и т. п.) не происходит, неизбежен вывод о том, что суммарно лесные сообщества имеют нулевой баланс: сколько они углекислого газа поглощают, столько его тут же и выделяют, следовательно, сколько они кислорода выделяют, столько его тут же и поглощают, леса ни в коей мере не являются лёгкими планеты. В коралловых же рифах ситуация иная: они связывают углекислый газ в виде карбонатов, из которых образуют многокилометровые толщи известняков. Конечно, коралловые рифы — не единственные лёгкие планеты. Лёгкими можно считать также болота (С02 связывается в торфе), озёра (С02 связывается в сапропеле или иных озёрных отложениях), сообщества рыхлых, особенно заиленных грунтов (С02 связывается как органическое вещество осадков, а также в известковых скелетных элементах, моллюсков, усоногих раков, фораминифер, полихет и др.), степные экосистемы (С02 связывается в мощном слое гумуса).
Высокая продуктивность коралловых рифов влияет и на прилегающие экосистемы. Непременной особенностью коралловых (как и любых других) рифов является шлейф детрита, сносимого с рифа, обогащающего донные сообщества. Кроме того, сезонные шторма выносят за пределы рифа много легкодоступной пищи. Это вызывает сезонные миграции нектона, связанные с этими периодическими событиями. Например, осенние шторма у северо-западного побережья Австралии вызывают миграции к коралловым рифам этого района планктоноядных рыб (сардин, китовых акул) и питающихся ими акул, тунцов и усатых китов.
Экосистемы коралловых рифов не только высокопродуктивны, но и хорошо сбалансированы, именно это позволяет им существовать в казалось бы бесплодных районах. Высокая сбалансированность обеспечивается высоким таксономическим разнообразием экосистем коралловых рифов, это не только одни из наиболее продуктивных сообществ Земли, но и одни из наиболее богатых видами, они обеспечивают существенную часть биоразнообразия морской биоты. Если бы коралловые рифы на Земле внезапно исчезли, то это бы привело к исчезновению до четверти, а, возможно, и более видов морской биоты. Чистый доход от эксплуатации экосистем коралловых рифов оценивают в настоящее время в $30 миллиардов в год.
Основу кораллового рифа, его скелет, образуют кораллы. Современные рифообразу- ющие кораллы относят примерно к 800 видам 110 родов. Большая их часть — мадрепоро- вые кораллы. Это — колониальные формы с мелкими полипами (размер одного полипа от долей мм до 1-2 см), колонии прирастают к твёрдому субстрату. Лишь мадрепоровые кораллы Fungiidae одиночные, свободнолежащие на рыхлых грунтах, размер их в поперечнике от 5 до 35 см и более, но Fungiidae не являются рифообразующими. Форма колоний рифообразующих кораллов видоспецифична и очень разнообразна, от массивных до весьма ажурных ветвящихся. Распределение колоний разной формы обусловлено в первую
![Распространение мелководных коралловых рифов. По Vernon (2000)]()
Рис. 3.56. Распространение мелководных коралловых рифов. По Vernon (2000).
очередь гидрологией: виды с массивными колониями живут на мелководьях внешней части рифа, где колонии других форм быстро бы разрушились из-за действия прибоя. Несмотря на пестроту и мозаичность распределения кораллов на рифе, основу сообщества образуют не более 10-15 видов мадрепоровых кораллов и 1-2 вида гидрокораллов. Первоначально рифообразующие кораллы называли герматипными (т. е. образующими биогермы — биогенные карбонатные постройки). Поскольку все мелководные рифообразующие кораллы содержат симбиотические зооксантеллы, этот термин стал применяться к кораллам с зооксантеллами. Это привело к некоторой путанице, поскольку эти свойства не жёстко связаны. Т. е. помимо рифообразующих кораллов с зооксантеллами, есть рифообразующие кораллы без зооксантелл, кораллы с зооксантеллами, не образующие рифы, и кораллы без зооксантелл, не образующие рифы. Ниже под герматипными кораллами будут подразумеваться, как это наиболее распространено в литературе, мелководные рифообразующие кораллы с симбиотическими зооксантеллами.
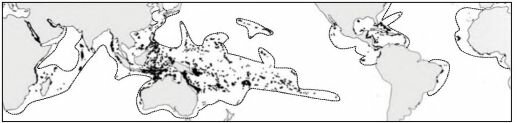
Мелководные коралловые рифы встречаются исключительно в водах, солёность которых не опускается ниже 30-31 %о. В приустьевых районах, где сказывается опресняющее действие рек, коралловые рифы угнетены или вовсе отсутствуют. Так, у устья Амазонки из-за её распресняющего действия коралловые рифы отсутствуют на протяжении почти 4 тыс. км (рис. 3.56, 3.60).
До сих пор широко распространено мнение о том, что распространение герматип- ных кораллов в высокие широты ограничивает температура воды. Карта, на которой проведена поверхностная изотерма в 20°С и показано распространение коралловых рифов имеется чуть ли не в каждой популярной книге о море. Наумов с соавторами (1985) даже указывают, что температура в местах образования коралловых рифов не опускается ниже 20,5°С, хотя герматипные кораллы способны существовать и при 18°С, но при этой температуре у них невозможен половой процесс (а, следовательно, и образование новых колоний), а рост сильно замедлен. В действительности, как указывает J.E.P. Veron (1995) это не совсем так. Так, из герматипных кораллов, обитающих у берегов Японии, 48% способны выносить 14,1°С, 22,5% — 10,4°С, а отдельные виды, например Oulastrea crispata, — и 0°С. Acropora hycicinthus у Японии является доминирующим видом в районах с минимальной среднемесячной температурой 15,3°С и смертность этого вида не возросла в 1980 г., когда температура упала до 13,7°С (Misaki, 1984, 1985). И, хотя распространение коралловых рифов и видов кораллов, образующих эти рифы, — не одно и то же (последнее гораздо шире), очевидно, что температура здесь не влияет прямо. Альтернативная гипотеза, которая находится в большем соответствии с имеющимися данными, объясняет положение границы распространения мелководных коралловых рифов в высоких широтах при низкой температуре взаимодействием их сообществ с сообществами макрофитов, в первую очередь бурых водорослей.
По направлению к высоким широтам происходит снижение числа видов герматипных кораллов. Какие-либо виды, специфические для сравнительно высокоширотных рифов, замещающие тропические, отсутствуют. Высокоширотные рифы образованы малым числом видов, случайно отобранных из небольшого числа эврибионтных видов с очень широким распространением. Сравнение видового состава отдельных районов разными индексами показывает сильное отличие периферических районов, что отражает случайный характер видового состава и малое число обитающих в сравниваемых районах видов, с биогеографической точки зрения эти районы не заслуживают выделения в самостоятельные регионы.
Герматипные кораллы всегда поселяются в местах, куда падает прямой солнечный свет. Существенной особенностью приэкваториальной области (где живут герматипные кораллы) является то, что солнце круглый год движется почти по одной траектории. В результате кораллы всегда находятся практически в одинаковых условиях. Несмотря на то, что вода в тропиках очень прозрачна, герматипные кораллы не встречаются глубже 80-100 м, а наибольшее их разнообразие наблюдается до глубины 15-25 м.
После образования скелета происходит заполнение пустот, превращение кораллового рифа в монолит. На этой стадии ведущая роль переходит к известковым водорослям, преимущественно красным (Corallinaceae, рода Porolithon и Lithotamnion), а также зелёным (Caulerpaceae, род Halimeda). Велико значение и других организмов с известковыми образованиями: фораминифер, губок, моллюсков (Tridacninae, Vermetidae), полихет. Скорость образования извести у водорослей гораздо выше, чем у кораллов. Именно они образуют основную массу карбонатной породы рифа. Живые кораллы имеют проективное покрытие не более 30-50% даже в зоне их активного роста, а часто lt;5%.
На небольших глубинах ресурсов пищи и света достаточно для всех герматипных кораллов, поэтому по этим факторам специализации нет. Она идёт по доступности субстрата, устойчивости к физическому стрессу (прибойность, мутность воды и т. п.) и выеданию, а также по стадиям сукцессии.
Географическое распространение склерактиний и коралловых рифов характеризуют следующие особенности:
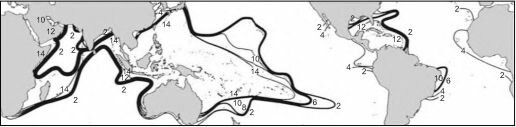
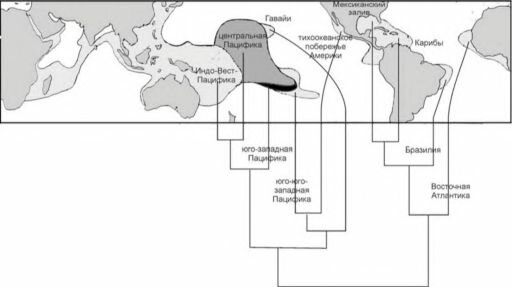
- По числу видов (рис. 3.57) резко выделяется Индо-Малайский центр видового разнообразия. Число обитающих здесь видов на порядок больше, чем в Карибском регионе и Атлантике вообще. Число видов в Карибском регионе примерно равно таковому в наименее разнообразных районах Индо-Пацифики.
- Разнообразие семейств (рис. 3.58) в Карибском и Индо-Пацифическом регионах одинаково, Индо-Малайский центр разнообразия не выражен. По-видимому, это объяснимо циркумтропическим распространением семейств склерактиний в эоцене и миоцене.
- На всех таксономических уровнях происходит отчётливое уменьшение разнообразия от низких к высоким широтам.
- Как число видов, так и семейств у восточных берегов Тихого и Атлантического океанов гораздо меньше, чем у западных.
Судя по отсутствию отличий на уровне семейств, атлантическая фауна склерактиний ранее могла быть столь же разнообразной, как и Индо-Пацифическая, но сейчас, по- видимому из-за эпизодических вымираний, она гораздо беднее.
Коралловые рифы Атлантики и Индо-Пацифики отличаются не только числом видов, но и таксономическим составом как самих склерактиний (рис. 3.60), так и сопутствующей биоты (табл. 3.2).
Предложено несколько объяснений высокого видового разнообразия Индо-Малайского архипелага и смежных районов:
- J. Briggs (1974, 1992, 1995) предположил, что этот район — центр современного происхождения новых видов, которые постепенно распространяются из него. Подтверждением этого предположения является увеличение среднего возраста родов кораллов по мере удаления от Индо-Вест Пацифики (рис. 3.59).
![Число видов мелководных рифообразующих кораллов в разных районах. По Vernon (2000)]()
Рис. 3.57. Число видов мелководных рифообразующих кораллов в разных районах. По Vernon (2000). ![Число семейств мелководных рифообразующих кораллов в разных районах. По Vernon]()
Рис. 3.58. Число семейств мелководных рифообразующих кораллов в разных районах. По Vernon
(2000). ![Средний возраст ныне живущих родов мелководных рифообразующих Scleractinia в млн. лет в разных районах]()
Рис. 3.59. Средний возраст ныне живущих родов мелководных рифообразующих Scleractinia в
млн. лет в разных районах.
Максимальный возраст рода был условно принят за 100 млн. лет, минимальный — 25 млн. лет. По Vernon (2000). ![Кладограмма сходства родового состава мелководных рифообразующих кораллов крупных районов Мирового океана]()
Рис. 3.60. Кладограмма сходства родового состава мелководных рифообразующих кораллов крупных районов Мирового океана.
В качестве меры сходства использован коэффициент различия Браун-Бланке. По Vernon (2000).
Таблица 3.2. Сравнение коралловых рифов Индо-Пацифики и Атлантики. По Наумову и др. (1985). Индо-Пацифика Атлантика
51
31
Gorgonaria
отсутствуют
600
=1200
=25000
число видов рифообразующих кораллов число родов рифообразующих кораллов общие 6 родов и, возможно, 1 вид второе место по значимости из Coelenterata Tubiporida
число видов коралловых рыб
число видов моллюсков
общее число видов, связанных с
сообществами мелководных коралловых рифов
Alcionaria иногда доминируют 2200 =5000
=125000
=500
80
- M.E.J. Wilson, B.R. Rosen (1998) изучили палеонтологические данные по мелководным рифообразующим кораллам и не нашли свидетельств наличия массовых коралловых рифов в этом районе до раннего миоцена (примерно 21 млн. лет назад). Из этого они сделали вывод о том, что здесь нет и не было центра происхождения (непонятно, почему они не допускают возможности возникновения центра происхождения позже, более того, коралловые рифы современный облик на Земле приобрели лишь в плиоцене - миоцене, в олигоцене же рифы образовывали также кораллиновые и зелёные водоросли, мшанки и некоторые фораминиферы, двустворки и гастроподы, роль склерактиний в построении рифов и их видовое разнообразие были сравнительно невелики). Причиной же современного высокого таксономического разнообразия этого региона они видят в том, что после сближения Австралии и Азии в этом районе образовалось большое количество в разной степени изолированных мелководных участков, степень изоляции которых к тому же сильно менялась во время оледенений. Это привело к возникновению большого числа новых таксонов в этих изолированных участках.
- J.F. Crame (1997) считает, что булыпее разнообразие тропических таксонов является не специфической особенностью самих тропических таксонов, а объясняется дли-
береговой риф
![]()
риф
барьерный риф
![]()
атолл
лагуна
![Типы коралловых рифов]()
Рис. 3.61. Типы коралловых рифов.
— R Jokiel и F.J. Martinelli (1992) построили математическую модель линий равномерно распределённых островов, помещённых в поверхностные течения, сходные с современными. Эта модель дала результат увеличения числа таксонов с востока на запад, т. е. тот же, что наблюдается сейчас.
Строение рифа определяет, в первую очередь, дифференциальный рост кораллов (рис. 3.61). Разрушение рифа в результате действия биотических (непосредственное выедание кораллов, разрушение скелета сверлильщиками) и абиотических (волновое
дополнили этот метод, учтя площадь доступную для образования рифов, и нашли, что только эти два фактора объясняют до 90% наблюдаемой долготной изменчивости таксономического разнообразия.
тельностью их существования, благодаря чему они имели больше времени для образования новых таксонов.
— R.F.G. Ormond и С.М. Roberts (1997)
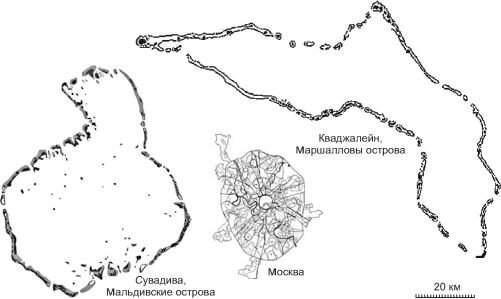
![Относительные размеры атоллов]()
Рис. 3.62. Относительные размеры атоллов.
воздействие) также вносит некоторый, но всё же не определяющий вклад. Различают три типа коралловых рифов: береговой, барьерный и атолл.
Береговой риф формируется на скальных плато небольших островов, лишённых заметного терригенного стока и не подверженных тектоническому опусканию. Характерная черта — отсутствие лагуны. Береговые рифы обычно невелики, но самый крупный береговой риф — береговой риф Красного моря достигает в длину 4 тыс. км.
Атоллы развиваются на вершинах подводных гор. Кольцевая их форма обусловлена появлением в центре рифа лагуны, когда:
- размеры рифа настолько велики, что в наиболее удалённых от открытого океана местах условия обитания кораллов ухудшаются из-за снижения турбулентности воды, в этих местах и образуется лагуна.
- снижение интенсивности водообмена приводит не только к снижению турблент- ности, но и к распреснению лагуны дождями, что дополнительно способствует её росту.
- в результате геологических процессов опускается основание рифа. Опускание компенсируется ростом кораллов, но растут они не везде с одинаковой скоростью, а преимущественно по наружному краю (где условия их обитания оптимальны).
Самые крупные атоллы — Сувадива (Мальдивские о-ва, Индийский океан) и Ква- джалейн, или Меньшикова (Маршалловы о-ва, Тихий океан). Лагуна атолла Меньшикова имеет 120 км в длину и до 30 км в ширину (рис. 3.62).
Отличительная особенность барьерного рифа — прибрежная лагуна — пространство открытой воды между рифом и берегом, где кораллы не растут. Лагуны барьерных рифов образуются в тех же случаях, что и лагуны атоллов, а также, когда имеется терриген- ный сток, угнетающий рост кораллов.
Наиболее крупные коралловые постройки принадлежат к барьерным рифам. Крупнейший барьерный риф (Большой Барьерный) расположен вдоль восточного побережья Австралии. Его протяжённость около 2000 км, ширина — от 15 до 300 км, а площадь — более 300 тыс. км2. Для сравнения: расстояние между Москвой и Чёрным морем почти в 2 раза меньше его длины, а площадь Московской области в 5 раз меньше.
Перечисленные выше типы — довольно абстрактны, реальные рифы обычно их совмещают. Так, Большой Барьерный риф в действительности является сложной системой из более, чем 2500 отдельных рифов, лагун, каналов и островов.
Как уже говорилось, герматипные кораллы в значительной степени существуют за счёт симбиотических водорослей. Эти водоросли принадлежат к разным семействам и даже классам, более того, имеются данные о возможности замены одного вида эндосимбионтов на другой у одной и той же колонии (неопубликованные данные Rowan и Knowlton, цит. по: Veron, 1995: 95). Кораллы с крупными полипами могут существовать исключительно за счёт хищного питания. Но булыпая часть кораллов в тени погибает за несколько месяцев. Только единичные виды герматипных кораллов способны факультативно существовать без симбиотических водорослей. Органическое вещество, синтезируемое симбионтами кораллов, потребляют кораллы. Развитие симбиоза с водорослями позволяет герматипным кораллам (как и другим группам с аналогичным симбиозом, существующим ныне и в прошлом) строить рифы и быть эдификаторами климаксных сообществ тропиков. Но и плата за это довольно велика:
- Герматипные кораллы способны обитать только на глубинах, где возможен достаточно эффективный фотосинтез.
- Сообщества коралловых рифов вступают в конкурентные взаимоотношения с сообществами макроводорослей, что ограничивает их распространение в высокие широты.
- В результате различных воздействий, возможно стрессового характера, герматипные кораллы способны утрачивать симбиотические водоросли. Это явление получило название «bleaching» — побеление (потерявшие симбиотические водоросли кораллы обесцвечиваются, белеют). Возможная адаптивная роль побеления — замена одного симбиотического вида водорослей на другой. Однако, поскольку без симбионтов герматипные кораллы существовать не могут, операция эта весьма рискованная. Существуют даже теории, объясняющие массовые вымирания склерактиний и других рифообразующих таксонов с симбиотическими водорослями разрушением симбиоза. Вне всяких сомнений, симбиоз с водорослями возникал и утрачивался неоднократно и независимо как у склерактиний, так и у других таксонов (рудисты, тридакны и т. п.). Однако слишком мало известно пока и о самом симбиозе, и о том, как и почему он возникает и исчезает, о происходящих при этом перестройках биохимии животных и растений.
Сообщество кораллового рифа потребляет энергию, синтезированную не только и, может быть, не столько герматипными кораллами, сколько другими фотосинтетиками сообщества. В первую очередь это не макрофиты и не известковые водоросли, о которых так много говорилось выше, а микрофиты, обрастающие обломки кораллов и интерстициальные формы. Количество эпифитона столь велико, что содержание органического углерода в обломках то же, что и в живых кораллах. Величина фотосинтеза также одного порядка на единицу площади. Но всё же без склерактиний существование экосистем коралловых рифов было бы невозможным, они — эдификаторы рифовых сообществ.
Более подробно с биологией мелководных коралловых рифов можно ознакомиться по книгам Д.В. Наумова с соавт. (1985), Ю.И. Сорокина (1990) и J.E.R Veron (1995).