Практика и традиции выделения сообществ макробентоса
Дно, в отличие от поверхности суши, длительное время было недоступно для визуального наблюдения. Изучение бентоса с судна традиционными орудиями лова можно сравнить с изучением суши с воздушного шара, плывущего над облаками.
Кто-то сказал, что с помощью трала в этом случае можно собрать мартовских котов и пару трубочистов. Исследователь-гидробиолог, в отличие от коллег, занимающихся изучением наземных экосистем, не имеет возможности отбирать пробы в конкретном сообществе, он может ориентироваться лишь весьма приблизительно, основываясь на данных по рельефу и материалам предыдущих съёмок.Первые исследования при описании бентоса выделяли не сообщества, а описывали отличия биоты разных грунтов. Например, в классическом труде по бентосу Чёрного моря, С.А. Зёрнов (1913) выделял биоценозы скал, песка, ракушечника, илистых берегов, мидиевого и фазеолинового ила и т. д. Такая практика выделения «сообществ» встречается и сейчас.
Постепенно было обнаружено, что в конкретном месте один из видов резко доминирует над остальными. Станции с доминированием одного вида стали относить к одному сообществу. Особенно широкое распространение практика выделения сообществ по видам-доминантам получила в отечественной гидробиологии (Броцкая, Зенкевич, 1939; Шо- рыгин, 1955 и др.). Этот метод выделения сообществ в изучении бентоса, как и в некоторых школах геоботаников, сохранилась до сих пор.
В исследованиях бентоса доминирование вида определяют в конкретной пробе или группе проб, взятых на одной и той же или соседних станциях (Броцкая и Зенкевич, 1939; Воробьев, 1949; Нейман, 1963). Доминирование следует оценивать по роли, которую играет вид в сообществе. Длительное время в качестве такого показателя использовали долю биомассы, часто с учетом частоты встречаемости. За рубежом часто вместо биомассы используют численность. Однако оба этих показателя имеют недостатки.
При выделении доминантов по биомассе, таковыми становятся крупные, но медленно растущие моллюски и иглокожие. При использовании численности в том же сообществе доминантом могутТаблица 3.1. Средние значения коэффициента удельной интенсивности метаболизма (с) при стандартной температуре 20°С, алометрического показателя и калорийности для некоторых групп макрозообентоса. По Алимову (1979); Кучеруку (1985); Голикову и др. (1988).
Таксон с d калорийность
ккал на 1 г
сырой массы
| Foraminifera | 0,3 | 0,75 | 0,3 |
| Spongia | 0,4 | 0,75 | 0,35 |
| Hydrozoa | 0,14 | 0,73 | 0,3 |
| Actiniaria, Alcionaria . . | 0,1 | 0,73 | 0,27 |
| Turbellaria | 0,84 | 0,81 | 0,87 |
| Nematoda | 0,83 | 0,8 | 0,86 |
| Polychaeta Errantia . . . | 0,74 | 0,78 | 0,85 |
| Polychaeta Sedentaria | 0,5 | 0,72 | 0,45 |
| Oligochaeta | 0,81 | 0,79 | 0,86 |
| Bryozoa | 0,14 | 0,7 | 0,3 |
| Brachiopda | 0,4 | 0,7 | 0,36 |
| Hydracarina | 0,7 | 0,75 | 0,4 |
| Copepoda | 0,97 | 0,78 | 0,7 |
| Mysida | 0,5 | 0,72 | 0,8 |
| Cirripedia | 0,14 | 0,69 | 0,3 |
| Amphipoda | 0,69 | 0,79 | 0,76 |
| Isopoda | 0,69 | 0,82 | 0,76 |
| Macrura | 1,1 | 0,87 | 0,82 |
| Anomura | 0,47 | 0,85 | 0,65 |
| Gastropoda | 0,42 | 0,71 | 0,35 |
| Bivalvia | 0,5 | 0,74 | 0,38 |
| в т.ч. Astartidae | 0,2 | 0,72 | 0,29 |
| Pectinidae | 0,7 | 0,8 | 0,5 |
| Mytilidae | 0,8 | 0,7 | 0,45 |
| Tellinidae | 0,33 | 0,71 | 0,4 |
| Asteroidea | 0,15 | 0,66 | 0,35 |
| Ophiuroidea | 0,15 | 0,72 | 0,3 |
| Holothuroidea | 0,15 | 0,72 | 0,28 |
| Tunicata | 0,1 | 0,6 | 0,25 |
| Pisces | 1,3 | 0,81 | 0,9 |
стать мелкие полихеты, имеющие низкую биомассу. Поскольку сообщество — функционирующая структура, правильнее для выделения доминирующих видов использовать не структурные показатели (численность, биомассу), а функциональные. В качестве такого показателя в последнее время широко используют поток энергии через популяции видов. Поток энергии можно считать пропорциональным интенсивности метаболизма популяции (трат на обмен). Интенсивность метаболизма популяции рассчитывают для животных по формуле:
R = c-NWd = cN1dBd,
где W — средний индивидуальный сырой вес особи, N — численность и В — биомасса на 1 м2, d — аллометрический показатель, С — специфичный для таксона коэффициент удельной интенсивности метаболизма.
При расчете потока энергии для макробентоса величину d обычно принимают равной 0,75, можно использовать значения из табл. 3.1; значения С специфичны для разных таксонов (табл. 3.1). В этой же таблице приведены средние величины удельной калорийности в ккал на 1 г сырой массы.
При оценке обилия раковинных моллюсков в значения сырого веса обычно вводят поправки на среднюю весовую долю их раковины, этот показатель колеблется в весьма широких пределах в зависимости от таксона, и его лучше определить самому для конкретного материала.
В качестве первого приближения можно использовать значения для Gastropoda — 0,5-0,67, Bivalvia — 0,9.Анализ имеющихся в литературе данных о продуктивности различных групп одноклеточных водорослей (морских и пресноводных), в основном диатомовых и перидиние- вых, показывает, что значения коэффициента d для этих организмов близки к 0,5 (Гутель- махер и др., 1980; Гутельмахер, 1983; Заика, 1983; Raven, Kubler, 2002).
Если резко доминирующего вида нет, то используют разные методы многомерного анализа.
После нанесения на карту сообществ, выделенных тем или иным способом, обнаруживается, что для булыпей части станций и сообществ характерны следующие особенности:
- станции, отнесённые к одному сообществу, расположены вполне закономерно;
- каждому сообществу свойственен свой тип гранулометрии;
- гранулометрические различия биотопов, близких по трофическому составу сообществ, заключаются в сортированности, т. е. степени преобладания и доли различных размерных частиц, при том, что преобладает одна и та же размерная фракция.
Таким образом, сообщество, выделенное по, казалось бы, формальным признакам, оказывается естественным образованием, обязанным своим возникновением различиям процесса седиментации и гидрологии, нередко очень тонким, а также, как будет показано в следующей главе, степенью его сукцессионной зрелости.
«Число, плозщадии контуры подобных биоценозов во многом условны и зависят от дроб- ностии размещения съёмочных станций» (Шунтов, 2001: 457).
В заключение необходимо отметить, что использование в качестве показателя роли вида в сообществе потока энергии, проходящего через его популяцию, хотя и стало привычным, вовсе не безупречно. В будущем несомненно произойдёт замена этого показателя на ещё более адекватный. В частности, идея о существовании вида-доминанта, определяющего сообщество, казалось бы многократно подтверждённая фактическими данными, не имеет теоретического обоснования. Виды, определяющие структуру и функционирование данного сообщества вовсе не обязаны доминировать в нём ни по численности, ни по биомассе, ни по любому другому аналогичному показателю.
Например, при любой методике отбора проб на ржаном поле, Homo sapiens не попадёт в виды-доминанты, более того, он вообще может не попасть в пробы, хотя именно он однозначно определяет структуру этого сообщества. Правильнее выделять сообщества по ключевым видам, или видам- эдификаторам (key-species), но универсальных методических приёмов этого не существует, такие виды сейчас устанавливают после изучения структуры и функционирования конкретных сообществ.С другой стороны, наиболее распространённая в данный момент практика — выделять сообщества на основе их физиономического сходства в данный момент — имеет и другие врождённые пороки. Мало того, что при этом фактически игнорируется сукцесси- онная динамика: более правильно выделять сообщества после её изучения, но в гидробиологии это очень отдалённая перспектива. Гораздо хуже, что физиономически сходные участки поверхности Земли относят к одному сообществу, хотя имеются многочисленные примеры того, что такое сходство может быть конвергентным.
Таким образом, на современном этапе сообщество — это, обычно, артефакт группировки данных, собранных на разных станциях (пробах). Иногда такие группировки удаётся интерпретировать биологически, иногда — нет.
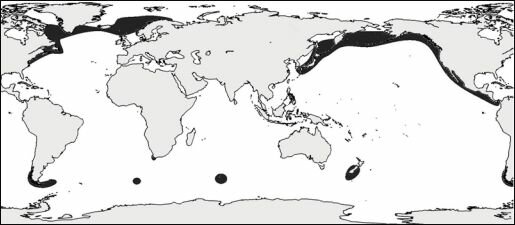
Рис. 3.7. Распространение сообществ, в которые входят разные виды морских ежей рода Brisaster.
По А.Н. Миронову (1989).
Еще по теме Практика и традиции выделения сообществ макробентоса:
- Таксономический состав и разнообразие макробентоса
- Продуктивность сообществ и жизненные стратегии водорослей. Морфофункциональный подход к изучению сообществ макрофитов
- СОСТАВ СООБЩЕСТВ И КОМПЛЕКСЫ СООБЩЕСТВ
- ЭКСПЕРТИЗА ВЕЩЕСТВЕННЫХ ДОКАЗАТЕЛЬСТВ В СУДЕБНО-СЛЕДСТВЕННОЙ ПРАКТИКЕ
- Биологическая концепция вида и практика систематики
- Эволюционное учение и практика сельского хозяйства
- Практика беспахотного земледелия: просто, быстро, эффективно
- Занятия по исследованию почвенных животных,рекомендуемые во время летней практики студентов
- Рифовые сообщества Особенности рифовых сообществ
- Метод выделения типов биот
- Сравнение разных методов выделения биогеографических регионов