Гидротермы и другие сообщества хемобиоса
Кроме фотоавтотрофов, в создании первичного органического вещества в море местами существенную роль играют хемоавтотрофы.
Открытие сообщества гидротерм — одно из крупнейших в биологии XX века, причём, в отличие от других открытий в биологии, например, в генетике, оно было совершенно неожиданным.
В недавно вышедшей монографии под редакцией А.В. Гебрука (2002) подробно рассматриваются все вопросы биологии гидротерм, изложенное ниже базируется, в основном, на ней.Впервые гидротермальные сообщества были обнаружены в мае 1976 года на Галапагосском рифте на глубине 2500 м. Судя по первым фотографиям, снятым там, для гид-
«горячий» источник
подушечные
лавы
покровные
лавы
морская
вода
~2°-3°С
глубокое (3-4 км)
гидротермальный раствор
гидротермальный раствор
1
1
| so42 -gt; s2 | НСОз -gt; со2, сн4 |
Mg2* Mn2* Са2* Fe2* Си2* Н2 |
|
|
_ \ 1 |
t t t t t | ||
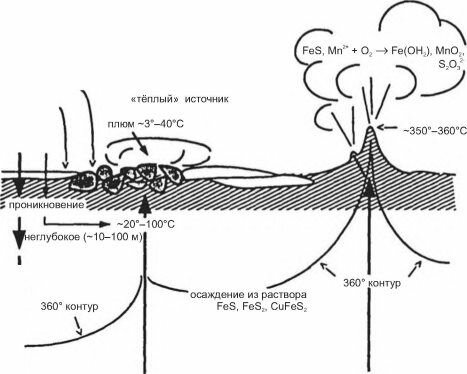
Рис. 3.71. Гидротермальная циркуляция. По А.В. Гебруку ( 2002).
ротерм характерны скопления крупных (до 20 см) белых двустворок, крупные крабы и банки из-под пива. Дальнейшие исследования показали, что в гидротермах происходит выход первичного вещества Земли. Точнее, происходит следующее (рис. 3.71, 3.72А, 3.73). По трещинам морская вода проникает в дно на глубину до нескольких сотен метров. Там она нагревается и насыщается газами, выделяющимися из магмы, в первую очередь сероводородом, а также водородом и ионами металлов (Mn, Fe и др.).
Нагретая вода устремляется вверх и выходит на поверхность в виде гейзера или более спокойных высачи- ваний (сипов). Температура воды на выходе из гейзеров достигает 350°С, но быстро падает, часто незначительно отличается от фоновой (рис. 3.72Б). Содержащиеся в воде сероводород, метан и другие газы окисляются, соответственно, сульфатредуцирующими, ме- танокисляющими и другими хемосинтезирующими бактериями.Создаваемое в гидротермах органическое вещество первично, поскольку синтезировано за счет энергии, полученной при окислении неорганических соединений, выделяю-
^вная зона хемосинтеза
H2S (S“,S203 ) -gt; Сорт + SO, NH/ (NOy) -gt; Copr. + N03 CH4 —^ Copr. + CO, хемоавтотрофы
симбиотрофы
бактериотрофы
V 6-23°C
хемоавтотрофы lt;
°сновная зона хемос^'
сообщество тёплых гидротерм
минизона хемосинтеза
[~хемоавтотрофы ^ 0имбио- и бактвпы
хемоавтотрофы
зона хемоси'
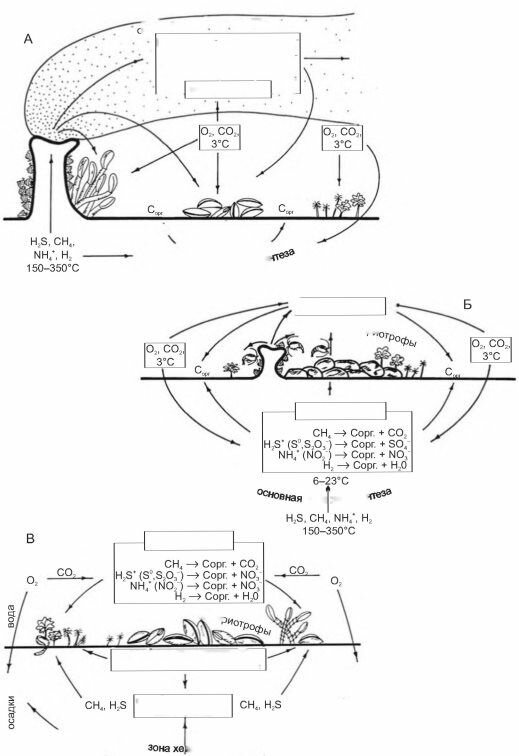
Рис. 3.72. Разные типы глубоководных хемот- рофных сообществ. По Гальченко (2002).
сообщество углеводородных сипов
зона хемосинтеза
хемоавтотрофы
симбио-и бактві
аэробные хемоавтотрофы СПН2„2 + 02—gt; Сорт + СО;
сульфатредукторы
матеногены
иотроф^и
щихся из магмы Земли. Гидротермы найдены во всех океанах. Лишь в Северном Ледовитом океане из-за его труднодоступности они пока не обнаружены, но, судя по тому, что там найдены створки Vesicomyidae — это вопрос времени.
Сходные сообщества образуются в сипах по окраинам континентов (рис. 3.72В). Здесь, в зонах субдукции осадки погружаются вместе с корой. В пассивных окраинах погружение осадков происходит в результате их накопления и соответствующего прогибания коры под их тяжестью (толщи осадков могут достигать 15 км и более). В обоих случаях погружение приводит к увеличению давления и росту температуры.
Это, в свою очередь, приводит к преобразованию органического вещества осадков с выделением в первую очередь метана, а также других газов. Первичная продукция здесь создаётся преимущественно за счет окисления метана.Третьим типом сообществ хемобиоса являются сообщества временных биотопов: крупные разлагающиеся органические остатки, такие как трупы китов и затонувшее судно, груженое консервированными ананасами, бобами, семенами подсолнечника и сизалем. Образующиеся здесь сообщества имеют похожий состав, включая даже отдельных представителей сообществ гидротерм, в том числе и вестиментифер.
Сообщества гидротерм, сипов и временных биотопов имеют много общих видов, они связаны генетически, образование этих сообществ и их распространение в океане могут быть поняты только при совместном их изучении, с биологической точки зрения между ними нет принципиальной разницы.
аномалия температуры
25-20°С 3-6°С 0°С
ВТ эутермальная олиготермальная фоновая
Н I 1 I 1 I
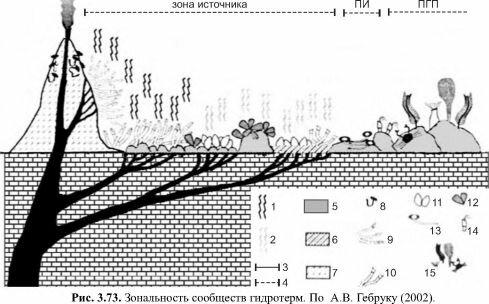
1 — диффузные истечения, визуально заметные по «муару»; 2 — низкотемпературные истечения, визуально не выраженные; 3 —термальные зоны (ВТ — высокотемпературная); 4 — зоны гидротермального поля (ПИ — периферия источника, ПГП — периферия гидротермального поля); 5 — базальты; 6 — рыхлые осадки; 7 — массивные сульфидные отложения; 8-15 — доминирующие фаунистические группировки: 8 — полихеты Alvinelli- dae, 9 — вестиментиферы Tevniida (кроме Escarpia), 10 — вестиментиферы Lamellibanchia и Escarpia, 11, 12 — двустворчатые моллюски Vesicomyidae (11) и Mytilidae (12), 13 — полихеты Serpulidae, 14 — усоногие раки, 15 — фоновая фауна.
Сообщества гидротерм, хотя и имеют много общих видов с сообществами сипов и временных биотопов, обладают ярко выраженными специфическими чертами, включая фауну, присущую только им. Эта фауна включает большое количество свойственных только ей таксонов высокого ранга: родов, семейств и даже подтипов.
Наиболее яркими представителями сообщества гидротерм являются вестиментиферы, помпейские черви — А1- vinellidae, двустворки Vesicomyidae и Mytilidae (Calliptogena). Эти организмы достигают весьма крупных размеров: наиболее крупная вестиментифера — Riftia — достигает 1,5 м в длину, помпейские черви достигают нескольких десятков см, Calliptogena — 20 см в длину и веса в 0,5 кг. Таксономический ранг специфичных таксонов — семейство — очень велик для морской биоты. Правда, ранг вестиментифер не вполне ясен. Нет сомнения, что это — один из таксонов погонофор. Но вот ранг погонофор — вопрос дискуссионный. Суммарная биомасса сообщества гидротерм составляет несколько кг/м2, что на 3-5 порядков превосходит биомассу соседних участков морского дна, не находящихся под влиянием гидротерм и одного порядка с биомассой наиболее продуктивных районов шельфа — шельфов умеренных широт. Высокую степень и ранг эндемизма принято интерпретировать как свидетельство большого возраста сообществ гидротерм, хотя, как показано в следующей главе, такое представление необоснованно. Однако наличие у эндемичных таксонов многочисленных глубоких приспособительных морфо-экологических модификаций, связанных в первую очередь с дыханием в условиях дефицита кислорода, по-видимому, свидетельствует о древности этих таксонов, хотя малое изменение генома симбиотических бактерий свидетельствует об обратном. О сравнительно небольшом возрасте некоторых таксонов (Alvinellidae, вестиментиферы), на мой взгляд, свидетельствует и незначительные размеры их ареала (они ещё не успели расселиться после возникновения). В последнее время всё большее признание получает мнение о том, что современные облигатные гидротермальные таксоны имеют кайнозойский возраст, а предположения об их более древнем возрасте слабо обоснованы (Миронов и др., 2002).Сами гидротермы — весьма древние сообщества. Для гидротермальных и других сообществ хемобиоса, современных и ископаемых, как и для сообществ высоких широт, характерно низкое разнообразие при высокой численности особей, входивших в сообщества видов.
Достоверные остатки гидротермальных сообществ, внешне сходных с современными, известны с силура. Эти сообщества также включали крупных червей в трубках (вестиментиферы?), мелких червей в трубках (Alvinellidae?) и крупных двустворок. Но существовали и другие сообщества, первичную продукцию в которых создавал микробный хемосинтез. Так, в раннем мелу существовали макробентосные сообщества фильтраторов эпифауны, которые, как и современные, находились в симбиотических отношениях с хе- мосинтетическими бактериями или непосредственно питались ими. Основой эти сообществ были трепостоматные мшанки, инкрустированные микробными оболочками, брахиоподы, черви в известковых трубках, сходные с вестиментиферами, конулярииды, сер- пулиды и остракоды (Невесская, 1999). В юре к организмам, связанным с бактериальным хемосинтезом относились двустворки-?люциниды (Gaillard, Rolin, 1986). Наиболее древние осадки формаций гидротермального происхождения имеют возраст 3,5-3,8 млрд. лет. Весьма вероятно, что на ранних этапах эволюции биосферы основным, а, возможно, и единственным источником энергии являлась не энергия Солнца, а энергия окисления восстановленных форм железа, марганца, серы и др.Сами сообщества гидротерм также различаются между собой. Прежде всего имеется зональность внутри отдельных гидротермальных источников в соответствии с градиентом абиотических условий, в первую очередь — концентрацией восстановленных соединений и первичной продукцией, формирующейся за счёт них (рис. 3.71). Имеются также данные об идущих в гидротермальных сообществах сукцессионных сменах. При расши-
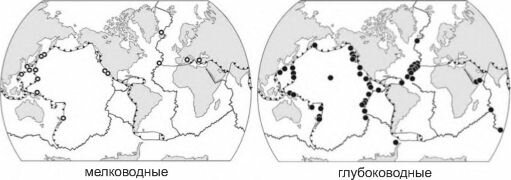
Рис. 3.74. Известные мелководные и глубоководные (только гидротермальные) сообщества хемобиоса. По В.Г. Тарасову и др. (2005).
фровке сукцессий существенные затруднения создаёт малая продолжительность существования самих гидротерм, сопоставимая с продолжительностью прохождения сукцессии.
В общем на основе незначительного числа наблюдений создаётся впечатление, что в восточной Пацифике вновь возникшие гидротермы колонизируют Alvinellidae, которых довольно быстро сменяют Riftia. Alvinellidae остаются только в наиболее высокотемпературном участке, где изменчивость условий, по-видимому, слишком высока. В свою очередь, Riftia постепенно вытесняют двустворчатые моллюски (в западной Пацифике — гастроподы). По мере продвижения сукцессии возрастает обилие и разнообразие десятиногих ракообразных. Эта общая картина в разных районах в той или иной степени варьирует. В любом месте колебания активности гидротермы способны вернуть сукцессию на любую из уже пройденных стадий.После обнаружения сообществ гидротерм, интерес к процессам хемосинтеза резко возрос и выяснилось, что как хемосинтез сам по себе, так и организмы, обеспечивающие себя энергией в той или иной степени за счёт симбиотических хемосинтетиков, распространены довольно широко и на малых глубинах от уреза воды (рис. 3.74).
Однако, несмотря на наличие в таких местах восстановленных соединений и активной бактериальной жизни, включая бактериальные маты, там не сформировалось сообществ, основным источником энергии которых был бы хемосинтез. Отмечают только сим- биотрофных Pogonophora и миксотрофных Bivalvia (Thyasiridae, Solemyidae, Lucinidae). Имеются также существенные различия в таксономическом составе сообществ разных глубин. В сообществах хемобиоса на глубинах менее 200 м нет специфичных для них видов макробентоса, а только виды, входящие в состав «нормальных» сообществ. Для более глубоководных сообществ (200-4100 м) описано около 400 специфических видов, специфические рода также появляются в сообществах глубже 200 м, семейства — глубже 386 м (Тарасов и др., 2005).
Причина столь резких различий мелководных и глубоководных сообществ, использующих хемоавтотрофию, по-видимому, состоит в том, что переход к использованию продукции хемосинтеза связан с целым рядом адаптаций: защита от токсинов, приспособленность к дефициту кислорода, расселению и поддержанию популяций в условиях эфемерных изолированных биотопов, а в случае гидротерм и к резким колебаниям температуры. Очевидно, что эти адаптации экологически осмысленны только при заметном преобладании потока энергии от хемосинтеза над фотосинтезом. Поэтому наиболее обоснованным выглядит мнение о том, что высокого потока энергии от хемосинтеза недостаточно для формирования хемотрофных сообществ. Необходимо также, чтобы поток энергии от фо-
Таблица 3.3. Основные отличия мелководных и глубоководных сообществ хемобиоса. По данным
В.Г. Тарасова и др. (2005).
черта мелководные глубоководные
глубина lt;200 м 200-4100 м
специфичные виды нет gt;400 м
специфичные рода нет есть, начиная с 200 м
специфичные семейства нет есть, начиная с 386 м
доминирующие виды
макробентоса неспецифичные специфичные
основной источник
энергии для сообщества фотосинтез хемосинтез
тосинтеза стал незначительным (Миронов и др., 2001, 2002). На меньших глубинах, даже в местах интенсивного поступления пригодных для хемосинтеза соединений (сульфидов, метана, восстановленных ионов металлов), продукция за счёт хемосинтеза не превышает продукции в этом же месте за счёт фотосинтеза. Поскольку площадь таких мест пренебре- жима мала по сравнению с местами с «нормальными» условиями, специализация на такие ресурсы бессмысленна. Следует также учесть, что в «нормальных» условиях хемо- (симбио)трофия также вполне обычное явление, поскольку в любом грунте на той или иной глубине появляется восстановленный слой. Хемоавтотрофные и хемосимбиотроф- ные (обычно — миксотрофные) организмы поэтому являются обычным компонентом «нормальных» бентосных сообществ, так что в местах повышенного потока восстановленных соединений происходит лишь увеличение их доли, т. е. перестройка уже имеющихся сообществ, обычно даже не очень существенная.
С увеличением глубины поток энергии от фотосинтеза уменьшается и, начиная с некоей глубины, становится энергетически выгодным возникновение сообществ, основным источником энергии для которых является хемосинтез. Глубина эта, вероятно, различна в разных местах, судя по имеющимся данным — это 200-400 м. Кроме того, от давления (которое есть линейная функция глубины) зависят растворимость газов, их парциальное давление, проницаемость мембран, равновесие химических реакций и т. п., возможно, что некоторые реакции, идущие при хемосинтезе, энергетически более выгодны при повышенном давлении, т. е. на булыпих глубинах. Так, имеются данные (de Angelis et al., 1991) о том, что физиологически достаточные скорости окисления метана требуют повышенного давления. Чем больше поток энергии, тем дальше заходит специализация сообществ, проявляющаяся во всё возрастающей специализации входящих в сообщества видов. Рост специализации, освоение нового адаптивного пространства приводит к возникновению специфических таксонов того или иного ранга.
Это различие мелководного и глубоководного биотопов определили и различия формирующихся в них сообществе (таблица 3.3), а также отсутствие интереса к мелководным сообществам с существенной долей хемосинтеза до тех пор, пока их изучение не потребовалось для лучшего понимания глубоководных хемоавтотрофных сообществ.
Суммарная площадь и продукция гидротерм и других сообществ, где органическое вещество создаётся хемосинтезом, на уровне Мирового океана пренебрежимо мала. Тем не менее это — древние и своеобразные ценозы, населенные специфичной биотой.
Биогеография гидротерм только начинает проясняться, многие районы (внетропиче- ское Южное полушарие, Северный Ледовитый и Индийский океаны) плохо или вовсе не изучены. Общая картина биогеографии гидротерм в современном представлении такова (рис. 3.75). Выделяют пять биогеографических районов. Наиболее разнообразны сообщества восточной Пацифики, наименее — Атлантики, где отсутствуют многие облигатные гидротермальные группы: вестиментиферы и Alvinellidae, тогда как креветки, доминирующие в Атлантике, играют в Пацифике вторые роли.
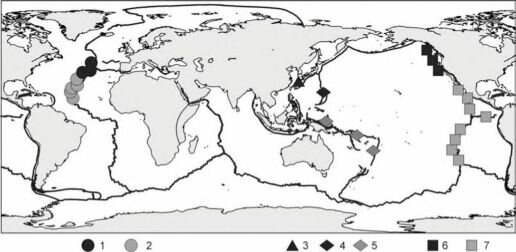
Рис. 3.75. Биогеографическое районирование гидротерм. По С.В. Галкину (2002).
Районы: 1,2 — атлантический (доминируют креветки Alvinocaridae и/или двустворки Mytilidae): 1 — североатлантический (батиальный), 2 — южноатлантический (абиссальный), 3 — южнояпонский (доминируют вести- ментиферы Lamellibranchia и двустворки Mytilidae); 4, 5 — западнотихоокеанский (доминируют гастроподы Provarmidae): 4 — западный, 5 — юго-западный; 6,7 — восточнотихоокеанский (доминируют вестиментиферы Tevniida и полихеты Alvinellidae): 6 — северовосточный, 7 — восточный-юговосточный.
Еще по теме Гидротермы и другие сообщества хемобиоса:
- Продуктивность сообществ и жизненные стратегии водорослей. Морфофункциональный подход к изучению сообществ макрофитов
- СОСТАВ СООБЩЕСТВ И КОМПЛЕКСЫ СООБЩЕСТВ
- Рифовые сообщества Особенности рифовых сообществ
- Гетеротрофные сообщества
- ДРУГИЕ ЭЛЕМЕНТЫ
- ДРУГИЕ ЭЛЕМЕНТЫ
- Другие письменные версии
- Другие тениидозы
- Другие овощи
- Другие двустворчатые и их промысловое значение
- Ассоциация и сообщество