Продуктивность сообществ и жизненные стратегии водорослей. Морфофункциональный подход к изучению сообществ макрофитов
Начиная разговор о первичной продукции, сначала коснёмся мифов, которые, как ни странно, бытуют даже среди биологов. «Большая биомасса» — совсем не значит «высокая продуктивность».
Фитопланктон, состоящий из множества микроскопических клеток, во много раз продуктивнее дубовой рощи. То же относится и к размерам отдельного организма: интенсивность обмена веществ у мелкого организма (например, у землеройки) гораздо выше, чем у крупного (слона)[******].Второй миф касается субъективного восприятия понятия «продукция»: автору этих строк довелось слышать доклад, в котором высочайшая продукция фитопланктона в эвт- рофированной бухте преподносилась как явление благоприятное и полезное, хотя «цветущая» вода в бухте дурно пахла и напоминала несвежий суп. Так явно положительный оттенок словосочетания «высокая урожайность» искусственных экосистем (полей и огородов) переносится и на «высокую продукцию» экосистем естественных. Собственно, самим экосистемам безразлично, чего в них много — дивных орхидей или токсичных цианобактерий. Не всё равно только человеку, которому предстоит жить среди или тех, или других. Проф. В.Н. Максимов, зав. кафедрой экологии Биофака МГУ, не устаёт повторять, что не надо охранять природу — она сама прекрасно справится, весь вопрос только в том, сможет ли человек существовать в этой новой природе, значит — охранять надо среду обитания человека. Такая постановка вопроса, на наш взгляд, гораздо более доходчива.
Эта преамбула имеет самое непосредственное отношение к тематике данного раздела. Продукция Мирового океана играет колоссальную роль в жизни человека. И речь не только о рыбе и прочих полезных гидробионтах — хотя их «урожайность» напрямую зависит от состояния среды их обитания. Прибрежные заросли макрофитов не только поставляют массу полезного сырья для пищевой, фармацевтической, лакокрасочной и даже мебельной промышленности (лучшими матрасами испокон веку считаются набитые сушёной морской травой).
Они обеспечивают приемлемое для человека и животных качество среды, обеспечивая аэрацию и самоочищение прибрежных вод. «Здоровые», сбалансированные сообщества, состоящие из многих компонентов, функционируют именно так, как нужно человеку, даже если он этого не понимает. Сообщества с нарушенным балансом элементов могут гораздо эффективнее использовать природные ресурсы, продуцировать гораздо больше фитомассы, но вот нужна ли такая фитомасса человеку — вопрос совсем не праздный. Мы уже упоминали «цветение» зелёной нитчатки Cladophora spp. в Анапской бухте, из-за чего знаменитый курорт теряет свою рекреационную привлекательность (Vershinin, Kamnev, 2001).Очевидно, что изучение закономерностей и механизмов формирования первичной продукции имеет исключительно большое теоретическое и практическое значение особенно в наше время, когда антропогенное загрязнение морей и океанов приобрело гло-
і і Laminaria, Канада, Атлантика
? ?dnnntradnn Macrocystis, Индийский океан
Thalassia, Индийский океан Spartina, Северная Америка, Атлантика
«-0*1 I I
•-о»!
I I I
I I I
«о
о
?
Thalassia, Карибское море
Macrocystis, Калифорния мангры, Пуэрто-Рико мангры, Флорида Zostera, Дания и штат Вашингтон Zostera, Аляска Spartina alterniflora прибрежный планктон 5 планктон открытого моря
4000
Валовая первичная продукция (total primary production) — это то количество органического вещества, которое образуется в процессе фотосинтеза на определённой площади за определённое время, или, по определению Р. Уиттекера (1980), та энергия, которая связана в этом веществе. В отличие от неё чистая первичная продукция (net primary production, net photosynthesis) — это то количество синтезированного органического вещества, которое пошло на строительство тела растения, за вычетом расходов на дыхание.
Для популяций или посадок однолетних растений биомасса может совпадать с продукцией: что выросло из семян или спор, т. е. практически с нуля, — то и есть и продукция (с момента начала роста), и биомасса (на данный момент). Для многолетников эти понятия (вернее, их численные выражения) совпадают только в первый вегетационный сезон. Далее остающиеся многолетние структуры (скелетные: стволы и ветки деревьев и кустов, стволики ламинарий, стволики и «ветви» фукоидов; или подземные — корни, корневища, ризомы и ризоиды) или совсем не вносят свой вклад в продукцию, или же он гораздо ниже, чем у ассимиляционных структур (листьев цветковых растений, листовых пластин многолетних водорослей). Чистая продукция проще всего определяется по разнице начальной и конечной биомассы популяции или массы одного растения.В старой советской литературе термин продуктивность трактовали как синоним термина «плодородие»: продуктивность водоёма = плодородие почвы, т. е. некое свойство того или иного биотопа или сообщества. На самом деле продуктивность — показатель динамический, он демонстрирует потенциальную способность растения, популяции или сообщества создавать продукцию на определённой площади в единицу времени. Короче говоря, авторам следует оговаривать, что они имеют в виду под тем или иным термином, тогда путаницы будет меньше.
Продуктивность донной растительности зависит от многих факторов: освещённости (вернее, фотосинтетически активной радиации — ФАР, лежащей в диапазоне 400-700 нм); наличия доступных питательных веществ; интенсивности гидродинамики, обеспечивающей приток этих веществ к талломам и т. д. Одним из наиболее важных факторов является форма самого таллома, которая определяет его способность использовать природные ресурсы, т.е. жизненную стратегию того или иного вида макрофитов. В сообществе совокупность растений строит сложную систему взаимоотношений со средой и друг с другом, причём основную роль здесь играет взаиморасположение растений, пространственное структурирование сообщества, обеспечивающее его оптимальное функционирование.
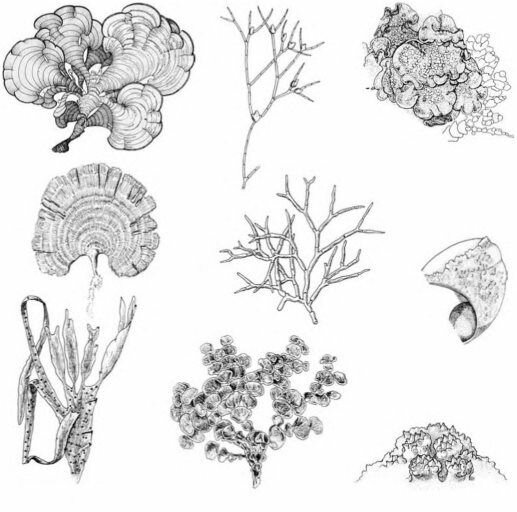
Морфо-функциональное направление, изучающее зависимость продукционных и иных характеристик от морфологии талломов отдельных видов и структурных особенностей фитоценоза и базирующееся на принципе точного соответствия структуры и функции, стало одним из основных направлений современной гидроботаники (Хайлов и др., 1992).Мы упоминали выше ряд систем жизненных форм водорослей (den Hartog, 1959; К. Петров, 2004), существуют и другие. Однако почти все они, базируясь на морфологии таллома, не учитывают адаптационное значение формы. Система, основанная на морфофункциональном подходе, была предложена М.М. Littler (1980) для исследования первичной продуктивности макрофитобентоса скалистых местообитаний тихоокеанского побережья Северной Америки. Были выделены следующие морфо-функциональные группы макроводорослей: 1) с талломом тонко-трубчатым или тонко-пластинчатым; 2) с нитчатым — тонким неразветвлённым или разветвлённым талломом; 3) с грубым разветвлённым талломом; 4) с талломом из толстых «ветвей» или пластин; 5) с членистым известковым и 6) с корковым, более или менее кальцинированным.
Наивысшие показатели первичной продукции демонстрируют почти исключительно зелёные и красные эфемероиды, а из бурых — лишь нитчатые формы, т. е. виды-оппор-
і s-'-:- а
бурая PetaIonia
красная Gigartina

красная Laurencia
зелёная Ulva ~
В ?+?--gt; •-
красная Cl Ас Porphyra
тонкопластинчатые
зелёная Codium
груборазветвлённые
ШООІпіЬйсь,
красная Ceramium
нитчатые или тонкоразветвлённые

зелёная Cladophora
красная Jania
бурая Padina
Mesophyllum lamellatum
зелёная Udotea
красная Amphiroa
Ftydrolithon на раковине
Neogoniolithon
красная
V Gracitaria зелёная Halimeda а также Laminaria,
см.
также рис. 3.36Sargassum, Fucus и др. (рис. 3.33, 3.35).
rfl
? о3
х га
га т 2 g X
ІІ1
4°о
плотные пластины и ветви
ЙЙйй
известковые с вертикальным талломом
корковые известковые багрянки
0lt;f #° xd
//ж
V
amp;
Ри с. 3.49. Морфо-функциональные группы водорослей. Данные по продукции по Littler et al. (1983), рисунки водорослей по Taylor (I960); Abbot, Hollenberg (1976); Rueness (1998).
тунисты. Ламинариевые, формирующие гигантские «морские леса», т. е. дающие колоссальную биомассу, по продуктивности сильно уступают этой мелочи. Таким образом, соотношение продукции и биомассы (Р/В-коэффициент) существенно выше у видов с небольшими, тонкими, примитивно устроенными талломами. Чистая продукция различается у перечисленных групп на порядки: для первой (Ulva, Petalonia, Porphyra) — 5,16 мг С/г в час (причём максимальные значения достигают 10-11); для второй (Cladophora, Chaetomorpha, Ceramium) — 2,47; для третьей (Codium, Laurencia, Gigartina) — 1,3; для четвёртой (Padina, Gracilaria, Sargas- sum, Fucus, Laminaria) — 0,76; для пятой (Coral- lina, Jania, Amphiroa) — 0,45 и для шестой (Neogoniolithon, Lithothamnion, Hydrolithon, Peyssonnelia) — всего лишь 0,07 мг С/г в час, что в 74 раза меньше, чем у первой группы (Littler, Arnold, 1982). Аналогичное исследование, проведённое в тропических водах (на барьерном рифе у побережья Белиза), дало несколько отличающиеся результаты: для первой группы — 5,06 мг С/г в час; для второй — 5,65; для третьей — 1,09; для четвёртой — 0,88; для пятой — 0,18 и для шестой — 0,02 мг С/г в час (Littler et al., 1983) (рис. 3.49). Несмотря на инверсию результатов в двух первых, самых продуктивных группах, основная тенденция осталась прежней, и даже полученные значения для групп близки, несмотря на разные океаны и климатические зоны.
выми шариками на цистозире (по: Хайлов и др., 1992).
Почему так получается? Дело в том, что продуктивность растений (и макроводорослей в том числе) связана с таким параметром, как удельная площадь поверхности (УПП) фотосинтезирующих органов — S/W, где S — площадь этих органов, a W — их масса. У водорослей, которые не только фотосинтезируют, но и поглощают питательные вещества поверхностью слоевища, эта зависимость еще больше, чем у высших растений. Наземные ботаники предпочитают обратный показатель W/S, называемый удельная поверхностная плотность. К.М. Хайлов (1988) на живых талломах ряда водорослей и на их гипсовых моделях показал, что параметр S/W имеет решающее влияние как на физико-химический обмен моделей, так и на углеродный обмен живых растений с окружающей их движущейся водой. При этом интенсификация обмена при постоянных условиях достигается ветвлением таллома.
Разработаны методики определения УПП, основанные на разделении слоевищ водорослей на цилиндрические и пластинчатые элементы, площадь поверхности которых определяется по известным формулам. Практически всё многообразие морфологии талломов можно свести к двум основным типам — осевому и пластинчатому, а также к разным их сочетаниям. Очевидно, что суммарная площадь поверхности сильно разветвлённых талломов выше, чем у разветвлённых слабо. Именно такими талломами обладает абсолютное большинство нитчаток — видов-оппортунистов. Пластинчатые талломы наращивают площадь своей поверхности за счёт рассечения пластины, образования на ней дочерних выростов — пролификаций, образования гофрировки, воланов, складчатости. Все эти структуры увеличивают площадь контакта таллома с водой и, соответственно, интенсивность обмена веществ, что подтверждено экспериментально в природных условиях (Хайлов и др., 1992).
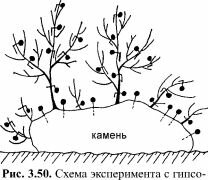
Методика таких натурных экспериментов не имеет аналогов в мировой практике и универсальна во всех случаях, когда нужно измерить локальное движение воды — в зарослях водорослей или вокруг единичного растения, на каких-либо конструкциях и т.д. Чтобы учесть интенсивность движения воды вокруг конкретных точек слоевища или субстрата, используют гипсовые шарики определённой массы (10-50 мг). Гипсовую смесь наносят на тонкие проволочки или синтетические нити, в два-три приёма наращивая шарики нужного размера (рис.3.50). На слоевище бурой Desmarestia (длиной до 60 см) удавалось разместить до 100 шариков, на слоевище красного Ceramium — 8-10 штук. Естественно, все шарики должны быть помечены — цветом или номерами. Скорость растворения гипса при определённых скоростях движения воды известна, поэтому по начальной (до эксперимента) и конечной (после эксперимента) массе шариков можно судить об интегральной подвижности воды во время экспозиции. Для той же цели используют и модели талломов, покрытые гипсовым слоем (Хайлов и др., 1988). Эта методика позволила выяснить, что вода гораздо активнее движется у разветвлений и неровностей талломов, а также у выступов и перегибов субстрата, образуя турбулентные потоки и обеспечивая постоянный подток питательных веществ. Именно поэтому на неровностях субстрата растительность всегда более богатая и мощная, чем на ровном дне, а разветвлённые талломы имеют более интенсивный обмен со средой и, соответственно, более продуктивны.
Площадь поверхности части таллома определяется с помощью простых геометрических и стереометрических формул: для осевых структур — по площади поверхности цилиндра, для пластинчатых — по площади поверхности той или иной геометрической фигуры или с использованием круглых высечек определённого диаметра (Миничева, 1987). Полученную величину относят к массе элемента слоевища и получают значение УПП элемента. Зная УПП отдельных элементов слоевища, можно вычислить общую площадь поверхности всего растения того или иного вида. Для этого сначала определяют общую поверхность Sk k-ой группы органов (листовых пластин, стволиков, ветвей определённого порядка и т.д.):
Sk = (S/W)k х Wk ,
где Wk — общая масса этой группы. Суммируя полученные величины по разным группам органов, получают общую поверхность таллома. Аналогично можно вычислить площадь поверхности размерно-весовых групп растений, популяции в целом, а имея представление о видовом составе и структуре доминирования конкретного сообщества — и площадь поверхности всего фитоценоза (Хайлов и др., 2004).
Следует учитывать, что величины УПП и общей поверхности талломов конкретного вида водоросли зависят от множества факторов: глубины произрастания, трофности среды, положения на элементах микрорельефа дна, а также от возраста самого таллома. Поэтому к предложенным числовым значениям УПП популяций различных видов (Миничева и др., 2003[††††††]) надо относиться с осторожностью. Так, УПП Phyllophora nervosa, определённая методом высечек, для северо-западного района Чёрного моря равна 14 м2/кг (Миничева, 1990), а для района Геленджика — всего 4 м2/кг (Рыбников, 1993а). Такие существенные различия могут объясняться тем, что авторы работали с разными экологическими формами филлофоры, известной своей морфологической изменчивостью.
Тем не менее, определённые величины УПП, даже если их численное выражение не точно и не абсолютно, дают представление о диапазоне УПП видов с разной морфологией. Например, у обильно разветвлённых тонких нитчаток Rhizoclonium implexum, Calli- thamnion corymbosum и Ectocarpus confervoides УПП равна 219 м2/кг, 165 м2/кг и 173 м2/кг, соответственно; у тонко-пластинчатых Ulva rigida и Porphyra leucosticta — 36 м2/кг и 63 м2/кг, соответственно; у морских высших растений — 10 м2/кг (Zostera marina) и 6,5 м2/кг (Potamogeton pectinatus). Для сравнения — УПП диатомей и цианобактерий достигают 3900-5800 м2/кг (Миничева и др., 2003). Возвращаясь к данным Littler, Arnold (1982), можно объяснить, почему водоросли с нитчатыми талломами оказались менее продуктивны, чем с пластинчатыми. Для представителей исследованных родов (нитчатки Ceramium, Chaetomorpha, Ciadophora) значения УПП не так уж высоки: 25-26 м2/кг у церамиума, 21-28 м2/кг у хетоморфы, 46-113 м2/кг у разных видов кладофоры (в среднем 79 м2/кг). То есть, значения УПП у этих нитчаток меньше или вполне сопоставимы с УПП тонко-пластинчатых талломов.
Таким образом, морфология тела растения играет первостепенную роль в его продуктивности. С ней непосредственно связаны стратегии выживания видов водорослей, выработанные в процессе эволюции. Характерные для нарушенных местообитаний (stressed communities) виды отличаются от видов, характерных для «здоровых» сообществ (nonstressed communities, late successional forms) по целому ряду признаков (Littler, Littler, 1980).
Оппортунисты (Opportunistic fomis)
- Быстрая колонизация освободившегося субстрата.
- В основном эфемеры и однолетники.
- Таллом примитивный, недифференцированный, с небольшой индивидуальной массой, но с высокой УПП (или с высоким отношением поверхности таллома к занимаемому объёму).
- Высокие скорость роста и чистая продукция, практически все части таллома фотосинтезируют.
- Высокая энергия размножения, почти все клетки таллома способны продуцировать репродуктивные структуры; образуется много пропагул с минимальными энергетическими затратами на производство каждой; плодоношение практически в течение всего года.
- Калорийность таллома высока и одинакова во всех его частях.
- Разные стадии жизненного цикла изоморфны, имеют одинаковые жизненные стратегии. Молодые талломы отличаются от взрослых только размером.
- Избегают фитофагов путём временной или пространственной непредсказуемости и быстро возобновляются за счёт быстрого роста.
Виды «здоровых» сообществ (Late Successional Forms).
- Занимают субстрат медленно, появляются в поздних стадиях сукцессии.
- Многолетники с более сложными и длительными жизненными циклами.
- Талломы сложно организованы, разделены на функционально различающиеся органы. Индивидуальная масса высока, но соотношение площади поверхности и занимаемого объёма низкое (как и УПП).
- Рост относительно медленный, чистая продукция невысока. Высокие затраты на дыхание нефотосинтезирующих органов и тканей.
- Относительно низкая энергия размножения, наличие специальных репродуктивных органов и тканей. Большие энергетические затраты на производство каждой пропагу- лы. Плодоношение сезонное.
- Калорийность таллома различается в разных его частях. Способность запасать вещества для переживания неблагоприятных сезонов.
- Разные стадии жизненного цикла могут иметь совершенно различные жизненные стратегии. Цикл в основном гетероморфный. Ювенильные талломы могут иметь стратегию выживания, аналогичную оппортунистам.
- Сложная структурная и химическая защита от фитофагов.
Оба типа жизненных стратегий имеют свои плюсы и минусы. Так, оппортунисты с примитивными, мелкими и обычно нежными талломами больше страдают от штормового воздействия, истирания льдом или песком; более высокие водоросли закрывают им свет; большая УПП способствует быстрому иссушению при отливах; они служат основной пищей фитофагам. Их пропагулы имеют очень высокую смертность. С другой стороны, большая поверхность позволяет эффективнее использовать питательные вещества, высокие скорости роста обеспечивают быстрое восстановление популяции, а отсутствие физиологической специализации обеспечивает им способность осваивать большое разнообразие биотопов.
Более высоко специализированные виды ограничены в выборе подходящих биотопов. В случае масштабных нарушений штормами или антропогенным воздействием они долго не восстанавливаются. Значительные энергетические затраты требуются на поддержание многолетних частей талломов, из-за этого снижен расход энергии на рост и размножение. Но их пропагулы обычно более жизнеспособны. Они успешно конкурируют за свет из-за наличия многолетних структур (стволики) и больших размеров. Структурная специализация разных элементов таллома и тканей обеспечивает более рациональное размещение талломов в пространстве, что позволяет им занимать лидирующие позиции в сообществе. Они устойчивее к внешним физическим воздействиям. Гетероморфная смена генераций способствует переживанию неблагоприятных сезонов и обеспечивает равновесные отношения с фитофагами (Littler, Littler, 1980).
Деление на две жизненные формы соответствует разобранным в предыдущей главе г- и К-стратегам в г-К-клаесификации или же видам-оппортунистам, или эксплерентам и виолентам в C-S-R классификации, и данная классификация имеет все их достоинства и недостатки (субъективность отнесения видов к той или иной группе, наличие переходных форм между двумя этими крайностями). Однако водоросли, представляющие эти крайние точки («opposite ends of the thallus-form spectrum», ibid., p.33), например, Ulva и Corallina, действительно очень сильно различаются по обозначенным параметрам и способностям. Так, на очищенном субстрате ульва образовала развитые заросли уже через 1,3 месяца, а кораллина появилась только через полгода. Интенсивность фотосинтеза ульвы была в 21 раз выше, чем у кораллины (3,78 и 0,18 мг С/г сухой массы в час). Калорийность таллома ульвы почти в полтора раза выше (5,44 и 3,86 ккал/г). При натурных экспериментах с фитофагами ульва за 48 часов потеряла более 43% популяции, а кораллина — менее 20%. Ульва имела самый низкий процент депигментированных тканей (0,6%), в то время как кораллина — самый высокий (81,7%), что свидетельствует о «стоимости» затрат на скелетные ткани у многолетников в ущерб фотосинтетической активности. После сильного шторма популяция ульвы была практически полностью — на 95,6% — разрушена, а популяция кораллины потеряла менее 30% талломов (заметим, что самой устойчивой оказалась ламинариевая Egregia — её популяция утратила всего 4,5% талломов). Сила прикрепления таллома у ульвы — самая низкая: чтобы оторвать таллом от субстрата требуется приложить усилие всего 0,6 кг/см2, для кораллины эта величина составляет 4,8 кг/см2, а для эгрегии — 28 кг/см2. При этом проростки эгрегии отрывались при приложении всего 13% от этого усилия (Littler, Littler, 1980).
Итак, морфо-функциональные особенности водорослей, проявляющиеся наиболее зримо в строении их талломов и в продуктивности, являются результатом отбора, который в разных случаях шёл разными путями. В часто нарушаемых местообитаниях преимущество получали виды-оппортунисты; в более спокойных и предсказуемых условиях К-от- бор создал виды, характерные для климаксных стадий сукцессии. При этом нельзя механически переносить на водоросли схемы, предложенные для наземной растительности. Макрофиты-оппортунисты имеют черты стресс-толерантов, сохраняя при этом высокие скорости роста и огромную энергию размножения. Водоросли, условно относимые к К- стратегам, образуют массу спор и гамет, что характерно для оппортунистов.
Как бы то ни было, морфологическое разнообразие макроводорослей позволяет им рационально осваивать жизненное пространство и оптимально использовать ресурсы, занимая все возможные ниши. Поэтому морфологию макрофитов нельзя рассматривать в отрыве от её функциональной нагрузки, прежде всего — эволюционно выработанного адаптивного значения. Строение слоевища водоросли, по выражению К.М. Хайлова с соавторами (1992: 224), «оптимизировано по отношению к потокам воды и к использованию химических ресурсов».
Следует отметить, что макроводоросли не только приспосабливаются к данным условиям среды, но некоторые из них способны активно менять её свойства. «Чемпионами» здесь являются известковые красные и зелёные водоросли, о роли которых в создании коралловых рифов и собственно водорослевых «построек» мы упоминали выше. Корковые известковые багрянки, имеющие самые низкие показатели продуктивности и скорости роста, «расплачиваются» этими потерями за невероятную живучесть и устойчивость как к влиянию фитофагов, так и к воздействию волн. Они способны «цементировать» даже подвижные грунты (галечник) так, что постепенно образуется сплошная корка, устойчивая к штормам.
Морфо-функциональные исследования позволяют не только понять закономерности функционирования прибрежных фитоценозов, но и правильно выбрать, например, объекты для культивирования. Например, при исследовании 8 штаммов агароносной багрянки Gracilaria tikvahiae оказалось, что они составляют непрерывный ряд по соотношению «площадь поверхности/объём» и, соответственно, продуктивность. Таким образом, можно выбрать именно те штаммы, свойства которых наилучшим образом подойдут к конкретным условиям культивирования. Где-то имеет смысл выбрать наиболее продуктивный, а где-то — менее продуктивный, но более устойчивый к внешним воздействиям (Hanisak et al., 1990).
Говоря о продуктивности макрофитов, нельзя не упомянуть их способность выделять в среду и поглощать из среды растворённое органическое вещество (РОВ). По некоторым оценкам, полученным при исследовании макрофитов Чёрного и Баренцева морей, годовая экскреция РОВ зелёными водорослями достигает 23% от их годовой валовой продукции, бурыми — 39% и красными — 38%. Значения экскреции зависят от сезона (в тёплое время года она выше) и от скорости роста талломов разных видов (у быстро растущих экскреция интенсивнее). При разрушении талломов в конце вегетационного сезона в воду в виде РОВ поступает ещё до 30% их продукции, т. е. общее поступление растворённой органики за год может составлять до 70% валовой продукции макрофитов (Khailov, Burlakova, 1969). У высших растений солёных маршей (Spartina alterniflora) эти значения ниже: 5-10% от чистой продукции надземных частей растения (Turner, 1976). Другие исследователи приводят ещё более скромные цифры, однако в любом случае — огромная масса прибрежной растительности продуцирует весьма значительное количество РОВ. Так, беломорский Fucus vesiculosus выделяет в воду 0,5-1 г углеводов и простых фенолов на 1 кг сухой массы в сутки. При биомассе фукоидов на Восточном Мурмане до 20 кг/м2 в воду ежесуточно выделяется до 2 г фенолов с квадратного метра (Камнев, 1989).
Макрофиты способны к органотрофии, т.е. поглощению РОВ из воды. Экспериментально показано, что, например, церамиум способен активно поглощать РОВ, меченое 14С, причём растущие апикальные части талломов поглощают его в 4 раза интенсивнее нижних частей. Возможно, за счёт поглощения РОВ в некоторой степени компенсируются потери роста и продукции, возникающие из-за взаимного затенения в плотных зарослях (Хайлов и др., 1992).
Поскольку поглощающая способность оппортунистов выше, чем у других видов, то они и получают преимущество в эвтрофированной среде. Состав фитоценоза меняется по мере приближения к источнику загрязнения. Например, исследования трёхкилометрового участка в районе выхода сточных вод (Чёрное море) показали, что на наибольшем удалении от него доминирует бурая многолетняя Cystoseira crinitci с небольшим количеством эпифитов; при приближении к источнику начинают появляться большие пятна Ulva и Enteromorpha, а на цистозире появляется всё большее количество эпифитов. У самого источника абсолютно доминирует энтероморфа. Т.е., по градиенту эвтрофикации снижается роль крупных многолетних видов, а численность мелких эфемеров и однолетников увеличивается (Празукин, 1989). В Балтийском море вытеснение сообществ Fucus vesiculosus сообществами нитчатых водорослей приобрело катастрофический характер в масштабах всего бассейна (Kangas et al., 1982; Kautsky et al., 1986; Eriksson et al., 2002 и многие другие). Общие закономерности процесса одинаковы везде: при эвтрофикации первыми из сообщества выпадают виды с минимальными значениями удельной площади поверхности, преимущество получают короткоживущие высокопродуктивные виды, общая площадь поверхности фитоценоза увеличивается (Миничева, 1996).
Повышенная трофность среды способна менять морфологию талломов и тех видов, которые изначально не приспособлены к высоким концентрациям биогенных элементов. Так, в конце 1980-х гг., когда эвтрофикация Чёрного моря приобрела глобальный характер, у Северокавказского побережья были обнаружены новые морфотипы прикреплённой Phyllophora nervosa — перистый (morphapennata) и курчавый (morpha crispa) (рис. 3.51). От изначальной morpha nervosa они отличались большим количеством пролификаций, за счёт которых площадь поверхности талломов сильно увеличивалась. Пролификации на «перистых» талломах были линейные, ланцетовидные или округлые, а на «курчавых» — сильно скрученные, очень тесно расположенные, из-за них таллом иногда приобретал почти цилиндрическую форму. Был прослежен полный переходный ряд от обычного таллома до курчавого через разную степень перистости, часто встречались «гибридные» талломы (рис. 3.52). В последние годы (с 1999 по 2007 гг.) в этом районе не были обнаружены «курчавые» талломы, но «перистые» сохранились. Видимо, «курчавая» форма была ответом на повысившееся содержание биогенных элементов, а затем она исчезла, может
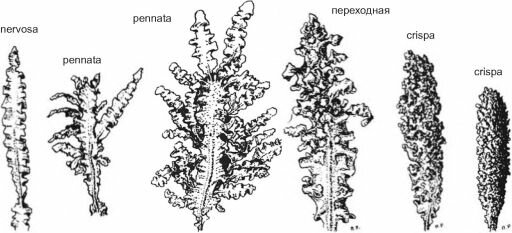
Рис. 3.51. Апикальные сегменты талломов филлофоры различных морфологических типов. Переходный ряд от morpha nervosa к morpha crispa в одном масштабе. Глубина до 1 м. По О.В. Максимовой, П.В. Рыбникову (1993).
«гибрид» nervosa и crispa
morpha pennata

Рис. 3.52. Морфотипы пргжреплённой Phyllophora nervosa. По О.В. Максимовой, П.В. Рыбникову (1993).
быть, из-за того, что её компактные талломы очень сильно обрастали мшанками, губками и прочим зооэпифитоном. А «перистая» филлофора оказалась более устойчивой; возможно, из-за интенсивного роста молодых пролификаций она более успешно сопротивлялась обрастанию.
Еще по теме Продуктивность сообществ и жизненные стратегии водорослей. Морфофункциональный подход к изучению сообществ макрофитов:
- Сообщества морских макрофитов[‡‡‡‡‡]
- Сообщества кораллиновых водорослей
- Сообщества ламинариевых водорослей
- Изучение структуры сообществ
- Сравнение структурно-функциональной организациимикробных сообществ различных природных зон:географический подход
- СОСТАВ СООБЩЕСТВ И КОМПЛЕКСЫ СООБЩЕСТВ
- МЕТОДЫ ИЗУЧЕНИЯ ВЗАИМООТНОШЕНИЙВ БИОТИЧЕСКОМ СООБЩЕСТВЕ
- Стратегии жизненных циклов
- Рифовые сообщества Особенности рифовых сообществ
- Классификации, рассматривающие три жизненные стратегии
- Гетеротрофные сообщества
- Ассоциация и сообщество
- Параллельные сообщества. Трофические зоны