ВЛИЯНИЕ ХОЛОДА НА РАСТЕНИЯ И ПРИСПОСОБЛЕНИЯ К НЕМУ
О пределе холода, который способны вынести растения в естественных условиях, дают представление величины предельно низких температур на земном шаре. Там, где зарегистрирована самая низкая температура (—90°С, станция «Восток» в Антарктиде) растительность отсутствует; а в районах, где живут растения, отмечена температура —68°С (Оймякон в Якутии, область таежных лесов из лиственницы — Larix dahurica).
Растительный покров обширных территорий земного шара (умеренные и арктические области, высокогорья) ежегодно в течение нескольких месяцев подвергается действию низких температур. Кроме того, в отдельных районах и в более теплые сезоны растения могут испытывать кратковременные воздействия пониженных температур (ночные и утренние заморозки). Наконец, есть местообитания, где вся жизнь растений проходит на весьма пониженном температурном фоне (арктические снежные и морские водоросли, приснежная — нивальная растительность в высокогорьях). Неудивительно, что естественный отбор выработал у растений ряд защитных приспособлений к неблагоприятному действию холода.
В чем опасность низких температур для растений? При отрицательных температурах замерзает вода вначале в межклетниках, затем внутри клеток, что ведет к обезвоживанию и механическому повреждению клеток, а затем к коагуляции белков и разрушению цитоплазмы. Но и в менее катастрофических случаях холод неблагоприятно сказывается на растениях, поскольку он тормозит основные физиологические процессы (рост, фотосинтез, образование хлорофилла, водообмен и т. д.), снижает энергетическую эффективность дыхания, резко замедляет скорость развития. Отрицательное влияние холода усиливается с увеличением продолжительности его действия. Возможны и механические повреждения: у древесных пород при продолжительных и сильных морозах образуются глубокие трещины в корке, коре и даже в древесине на стволах и крупных ветвях (морозобоины, или морозобойные трещины).
Кроме непосредственного влияния низкой температуры на растения под действием холода возникают и другие неблагоприятные явления. Например, уплотнение и растрескивание замерзшей почвы приводит к разрыву и механическому повреждению корней, образование ледяной корки на поверхности почвы ухудшает аэрацию и дыхание корней. Под толстым и долго лежащим снежным покровом при температуре около 0°С наблюдается зимнее «выпре- вание», истощение и гибель растений в связи с расходом резервных веществ на дыхание, грибными заболеваниями («снежная плесень») и т. д., а в случае избыточно увлажненной почвы для растений опасно также зимнее «вымокание». В тундре и северной тайге распространено явление морозного «выпирания» растений, которое вызывается неравномерным замерзанием и расширением почвенной влаги. При этом возникают силы, выталкивающие растение из почвы, в результате чего происходит «выпучивание» целых дернин, оголение и обрывы корней и т. д. вплоть до повала небольших деревьев. Поэтому кроме собственно холодостойкости (или морозостойкости) — способности переносить прямое действие низких температур, различают еще зимостойкость растений — способность к перенесению всех перечисленных выше неблагоприятных зимних условий.
Особо следует остановиться на том, как влияет на растения низкая температура почры. Холодные почвы в сочетании с умереннотеплым режимом воздушной среды растений (а иногда и со значительным нагреванием надземных частей растений) — явление нередкое. Таковы условия жизни растений на болотах (см. с. 196) и заболоченных лугах с тяжелыми почвами, в некоторых тундровых и высокогорных местообитаниях и в обширных областях вечной мерзлоты (около 20% всей суши), где в период вегетации оттаивает лишь неглубокий, так называемый «деятельный» слой почвы. В условиях пониженных температур почвы после снеготаяния (0—10°С) проходит значительная часть вегетации ранневесенних лесных растений — «подснежников». Наконец, кратковременные периоды резкого несоответствия холодных почв и прогретого воздуха испытывают ранней весной многие растения умеренного климата (в том числе и древесные породы).
Еще в прошлом веке немецкий физиолог Ю. Сакс показал, что при охлаждении почвы до околонулевых температур (обкладывание горшка льдом) может наступить завядание даже обильно политых растений, поскольку при низких температурах корни не способны интенсивно всасывать воду. На этом основании в экологии распространилось мнение о «физиологической сухости» местообитаний с холодными почвами (т. е. недоступности влаги растениям при ее физическом обилии). При этом упускали из виду, что Сакс и другие физиологи свои опыты производили с достаточно теплолюбивыми растениями (огурцы, тыква, салат и др.) и что в природных холодных местообитаниях растения, для которых низкие температуры почв служат естественным фоном, возможно, реагируют на них совсем иначе. Действительно, современные исследования показали, что у большинства растений тундр, болот, у ранневесенних лесных эфемероидов отсутствуют те явления угнетения (затруднение всасывания воды, расстройства водного режима и т. д.), которые могли бы быть вызваны «физиологической сухостью» холодных почв. Это же показано и для многих растений в областях вечной мерзлоты. Вместе с тем нельзя полностью отрицать угнетающее влияние низких температур на всасывание влаги и другие стороны жизнедеятельности корней (дыхание, рост и др.), а также на активность почвенной микрофлоры. Оно без сомнения имеет значение в комплексе трудных условий для жизни растений в холодных местообитаниях. «Физиологическая сухость», «физиологическая засуха» из-за низкой температуры почв возможны в жизни растений в наиболее трудных условиях, например при выращивании на холодных почвах теплолюбивых растений или ранней весной для древесных пород, когда еще необлиственные ветви сильно нагреваются (до 30—35°С) и увеличивают потерю влаги, а интенсивная работа корневых систем еще не началась.
Каких-либо специальных морфологических приспособлений, защищающих от холода, у растений нет, скорее можно говорить о за-
![Розетка листьев древовидного растения Lobelia keniensis (высокогорья Кении, 4500 м, Восточная Африка) (по Hedberg О., 1964).<div class=]()
А —днем, Б — ночью" />
Рис. 63. Розетка листьев древовидного растения Lobelia keniensis (высокогорья Кении, 4500 м, Восточная Африка) (по Hedberg О., 1964). А —днем, Б — ночью
щите от всего комплекса неблагоприятных условий в холодных местообитаниях, включающего сильные ветры, возможность иссушения и т. д. У растений холодных областей (или у переносящих холодные зимы) часто встречаются такие защитные морфологические особенности, как опушение почечных чешуй, зимнее засмоление почек (у хвойных), утолщенный пробковый слой, толстая кутикула, опушение листьев и т. д. Однако их защитное действие имело бы смысл лишь для сохранения собственного тепла гомеотермных организмов, для растений же эти черты, хотя и способствуют терморегуляции (уменьшение лучеиспускания), в основном важны как защита от иссушения. В растительном мире есть интересные примеры адаптаций, направленных на сохранение (хотя и кратковременное) тепла в отдельных частях растения. В высокогорьях Восточной Африки и Южной Америки у гигантских «розеточных» деревьев из родов Senecio, Lobelia, Espeletia и других от частых ночных морозов существует такая защита: ночью листья розетки закрываются, защищая наиболее уязвимые части — растущие верхушки (рис. 63). У некоторых видов листья опушены снаружи, у других в розетке скапливается выделяемая растением вода; ночью замерзает лишь ее поверхностный слой, а конусы нарастания оказываются защищенными от мороза в своеобразной «ванне».
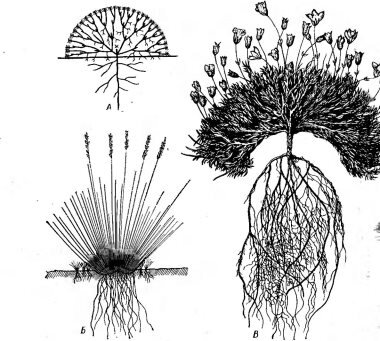
Среди морфологических адаптаций растений к жизни в холодных местообитаниях важное значение имеют небольшие размеры и особые формы роста. Не только многие травянистые многолетни-
![Нанизм высокогорных растений. А — В — растения альпийского пояса Западного Кавказа, 3000—3500 м над уровнем моря (фото Примы В. М.). lt;4 —Jurinea; filicijotia; Б—Valdivia jormosa, В — Arabis caucasica; Г — карликовая ива — Salix]()
Рис. 64 Нанизм высокогорных растений.
А — В — растения альпийского пояса Западного Кавказа, 3000—3500 м над уровнем моря (фото Примы В. М.). lt;4 —Jurinea; filicijotia; Б—Valdivia jormosa, В — Arabis caucasica; Г — карликовая ива — Salix her-
bacea (Восточные Карпаты, 1600 м над уровнем моря)
![]()
![]()
![Продолжение]()
Рис. 64. Продолжение
ки, но также кустарники и кустарнички полярных и высокогорных областей имеют высоту не более нескольких сантиметров, сильно сближенные междоузлия, очень мелкие листья (явление нанизма, или карликовости) (рис. 64). Кроме хорошо известного примера — карликовой березки (Betula папа), можно назвать карликовые ивы (Salix polaris, S. arctica, S. herbacea) и многие другие. Обычно высота этих растений соответствует глубине снежного покрова, под которым зимуют растения, так как все части, выступающие над снегом, гибнут от замерзания и высыхания. Очевидно, в образовании карликовых форм в холодных местообитаниях немалую роль играют и бедность почвенного питания в результате подавления активности микробов, и торможение фотосинтеза низкими температурами. Но независимо от способа образования карликовые формы дают известное преимущество растениям в приспособлении к низким температурам: они располагаются в припочвенной экологической микронише, наиболее прогреваемой летом (см. рис. 51), а зимой хорошо защищены снежным покровом и получают дополнительный (хотя и небольшой) приток тепла из глубины почвы.
Другая адаптивная особенность формы роста — переход сравнительно крупных растений (кустарников и даже деревьев) от орто- тропного (вертикального) к плагиотропному (горизонтальному) росту и образование стелющихся форм — стланцев, стлаников, стланичков. Такие формы способны образовывать кедровый стланик (Pinus pumila), можжевельник (Juniperus sibirica, J.
сот- munis, J, turkestanica), рябина и др. Ветви стланцев распластаны по земле и приподнимаются не выше обычной глубины снежного покрова. Иногда это результат отмирания ствола и разрастания нижних ветвей (например у ели, рис. 65), иногда это рост дерева как бы «лежа на боку» с плагиотропным, укоренившимся во многих
![Ветвление ели и переход к стланиковой форме в местообитаниях с сильными ветрами на верхней границе леса (Высокие Татры, Чехословакия)]()
Рис. 65. Ветвление ели и переход к стланиковой форме в местообитаниях с сильными ветрами на верхней границе леса (Высокие Татры, Чехословакия)
(по Plesnik Р., 1971)
местах стволом и приподнимающимися ветвями (кедровый стланик, рис. 66). Интересная особенность некоторых древесных и кустарниковых стлаников — постоянное отмирание старой части ствола и нарастание «верхушки», в результате чего трудно определить возраст особи.
Стланики распространены в высокогорных и полярных областях, в условиях, которых уже не выдерживают древесные породы (например, на верхней границе леса). Своеобразные «стланиковые» формы в крайних условиях встречаются и у кустарничков (рис. 67), и даже у видов лишайников, обычно имеющих прямостоячий кустистый рост: на скалах Антарктиды они образуют стелющиеся слоевища,
Рис. 66. Кедровый стланик — Pinus pumila (по Тихомирову Б. А. из Шен-
никова А. П., 1950)
![]()
В зависимости от условий возможны видоизменения роста одного и того же вида (рис. 68). Но есть виды, целиком перешедшие к форме стланика, например горный сосновый стланик, произрастающий в Альпах и Карпатах — Pinus mughus, выделенный в качестве самостоятельного вида из сосны горной — Pinus montana.
![]()
Н 8
К числу форм роста, способствующих выживанию растений в холодных местообитаниях, принадлежит еще одна чрезвычайно
![Можжевельник туркестанский — Juniperus turkestanica на склонах хребта Терскей-Алатау (по Серебрякову И. Г., 1955)]()
Рис. 68. Можжевельник туркестанский — Juniperus turkestanica на склонах хребта Терскей-Алатау (по Серебрякову И. Г., 1955). А — древовидная форма (лугово-лесной пояс, 2900 м над уровнем моря); Б — стланик (субальпийский пояс, 3200 м над уровнем моря)
своеобразная — подушковидная. Форма растения-подушки (рис. 69) образуется в результате усиленного ветвления и крайне замедленного роста скелетных осей и побегов. Мелкие ксерофильные листья и цветки расположены по периферии подушки. Между отдельными ветвями скапливаются мелкозем, пыль, мелкие камни. В результате некоторые виды растений-подушек приобретают большую компактность и необычайную плотность: по таким растениям можно ходить, как по твердой почве. Таковы Silene acaalis. Gypsophila aretioides, Androsace helvetica, Acantholimon diapensioides. Издали их трудно отличить от валунов. Менее плотны колючие подушки из родов Eurotia, Saxifraga.
Растения-подушки бывают разных размеров (до 1 м в поперечнике) и разнообразных очертаний: полушаровидные, плоские, вогнутые, иногда довольно причудливых форм (в Австралии и Новой Зеландии их называют «растительными овцами»).
Благодаря компактной структуре растения-подушки успешно противостоят холодным ветрам. Поверхность их нагревается почти так же, как и поверхность почвы, а колебания температуры внутри менее выражены, чем в окружающей среде. Отмечены случаи значительного повышения температуры внутри подушки; например, у наиболее распространенного вида высокогорий Центрального Тянь- Шаня Dryadanthe tetrandra при температуре воздуха 10°С внутри
![Растения-подушки (по Кернеру А., 1896; Заленскому О. В., 1948; Тихомирову Б. А., 1963; Strasburger Е.. et al., 1962; Hedberg О., 1964)]()
ikill ( .ivLu іг.л
Рис. 69 Растения-подушки (по Кернеру А., 1896; Заленскому О. В., 1948; Тихомирову Б. А., 1963; Strasburger Е.. et al., 1962; Hedberg О., 1964). А — схема ветвления растения-подушки; Б — высокогорный африканский злак — Festuca pilgeri с подушкообразной дерниной; В —арктическое растение-по
душка — Minuartia' arctica; Г — высокогорное памирское растение-подушка — Acantholimon diapensioides; Д — гигантские подушки («растения-овцы») в горах Австралии
подушки температура доходила до 23°С благодаря аккумуляции тепла в этом своеобразном «парнике». В связи с медленным ростом растения-подушки по долговечности вполне сравнимы с деревьями. Так, на Памире подушка Acantholimoti hedini диаметром 3 см имела возраст 10—12 лет, при 10 см — 30—35 лет, а возраст крупных лодушек достигал не одной сотни лет.
В пределах общей формы растений-подушек существует экологическое разнообразие: например в горах, окружающих Средиземное море, распространены менее компактные по строению ксе- рофильные «колючие подушки», которые не встречаются высоко в горах, так как малоустойчивы к холоду, но зато очень устойчивы к засухе. Рыхлое строение подушки здесь оказывается более выгодным для растения, чем компактное, так как в условиях летней засухи и сильной инсоляции снижает опасность перегрева ее поверхности. Температура поверхности средиземноморских подушек обычно ниже температуры воздуха благодаря сильной транспирации, а внутри подушки создается особый микроклимат; например, влажность воздуха держится на уровне 70—80% при влажности наружного воздуха 30%. Таким образом, здесь форма подушки — это приспособление к совсем иному комплексу факторов, отсюда и ее иная «конструкция».
Среди других особенностей роста, помогающих растениям преодолевать действие холода, следует еще упомянуть различные приспособления, направленные на углубление зимующих частей растений в почву. Это развитие контрактильных (сократительных) корней — толстых и мясистых, с сильноразвитой механической тканью. Осенью они высыхают и сильно сокращаются в длину (что хорошо заметно по поперечной морщинистости), при этом возникают силы, втягивающие в почву зимующие почки возобновления, луковицы, корни, корневища (рис. 70).
Контрактильные корни встречаются у многих растений высокогорий, тундр и других холодных местообитаний. Они позволяют, в частности, успешно противостоять морозному выпиранию растений из почвы. В последнем случае они не только втягивают почку возобновления, но и ориентируют ее перпендикулярно поверхности, если растение повалено. Глубина втягивания контрактильными корнями варьирует от сантиметра до нескольких десятков сантиметров в зависимости от особенностей растения и механического состава почвы.
Адаптивное изменение формы как защита от холода — явление, ограниченное в основном холодными районами. Между тем действие холода испытывают и растения более умеренных областей. Гораздо более универсальны физиологические способы защиты. Они направлены прежде всего на снижение точки замерзания клеточного сока, предохранение воды от вымерзания и т. д. Отсюда такие особенности холодостойких растений, как повышение концентрации клеточного сока, главным образом за счет растворимых углеводов. Известно, что при осеннем повышении холодостойкости («закаливании») крахмал превращается в растворимые сахара. Другая черта
Рис. 70. Контрактильные корни. А — Arum maculatum (углубление в почву от прорастания семени до взрослого растения); Б — участок корня Lilium martagon с поперечными морщинами (по Strasburger Е. et al., 1962);
В — луковицы Scilla sibirica
холодостойких растений — повышение доли коллоидно-связанной воды в общем водном запасе.
При медленном снижении температуры растения могут вынести охлаждение ниже точки замерзания клеточного сока в состоянии переохлаждения (без образования льда). Как показывают эксперименты, уровень точек переохлаждения и замерзания тесно связан с
![Точки переохлаждения и замерзания у дубравных растений. А — ранневесенние эфемероиды; Б — летневегетирующие виды:]()
А Б
I 2 | З | 4 | 5 | S | 7
1 ’і L п II р
I I
Рис. 71. Точки переохлаждения и замерзания у дубравных растений. А — ранневесенние эфемероиды; Б — летневегетирующие виды:
1 — Scilla sibirica, 2 — Corydalis halleri, 3 — Gagea lutea, 4 — Ficaria verna, 5 — Aego- podium podagraria, 6 — Asarum europaeum, 7 — Stellaria holostea. a — подземные органы, б — листья. На каждой вертикальной линии нижняя точка — температура переохлаждения, верхняя — температура замерзания
температурными условиями обитания (рис. 71). Однако у растений состояние переохлаждения возможно лишь при небольшом холоде (несколько градусов ниже нуля). Гораздо более действенным этот путь адаптации оказывается у других пойкилотермных организмов— насекомых, у которых роль антифризов играют глицерин, трегалоза и другие защитные вещества (открыто зимующие насекомые могут вынести переохлаждение клеточных соков без замерзания до —30°С).
Многие растения способны сохранять жизнеспособность и в промерзшем состоянии. Есть виды, замерзающие осенью в фазе цветения и продолжающие цвести после оттаивания весной (мокрица — Stellaria media, маргаритка — Beilis perennis, арктический хрен — Cochlearia fenestrata и др.). Ранневесенние лесные эфемероиды («подснежники») в течение короткой вегетации неоднократно переносят весенние ночные заморозки: цветки и листья промерзают до стекловидно-хрупкого состояния и покрываются инеем, но уже через 2—3 ч после восхода солнца оттаивают и возвращаются в обычное состояние. Хорошо известна способность мхов и лишайников переносить длительное промерзание зимой в состоянии анабиоза. В одном из опытов лишайник Cladonia замораживали при —15°С на 110 недель (более двух лет!). После оттаивания лишайник оказался живым и вполне жизнеспособным, у него возобновились фотосинтез и рост. Очевидно, у лишайников в крайне холодных условиях существования периоды такого анабиоза очень длительны, а рост и активная жизнедеятельность осуществляются лишь в короткие благоприятные периоды (причем не каждый год). Такое частое прерывание активной жизни на долгие сроки, по-видимому, объясняет колоссальный возраст многих лишайников, определенный радиоуглеродным методом (до 1300 лет у Rhizocarpon geographicum в Альпах, до 4500 лет у лишайников в Западной Гренландии).
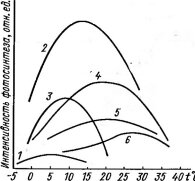
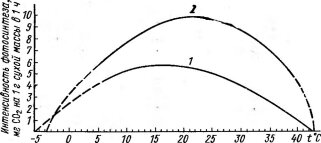
Рис. 72. Зависимость фотосинтеза от температуры у растений различных по тепловому режиму местообитаний (по данным разных авторов, схематизировано):
/ — субарктический лишайник — Nephroma arcticum (Сев. Финляндия), 2 — осока — Carex melanostachya (высокогорная криофильная пустошь, Гиссар- ский хребет, Таджикистан), 3—арктический вид — Oxytropis tschuktschorum (о, Врангеля), 4—ранневесенний эфемероид — Leticojum vertium (Австрия), 5 — лиственница — Larix decidua (Австрия), 6 — лимои — Citrus limonum (культура с зимней защитой от морозов)
![]()
Анабиоз — «крайняя мера» в борьбе растения с холодом, приводящая к приостановке жизненных процессов и резкому снижению продуктивности. Гораздо большее значение в адаптации растений к холоду имеет возможность сохранения нормальной жизнедеятельности путем снижения температурных оптимумов физиологических процессов и нижних температурных границ, при которых эти процессы возможны. Как видно на примере оптимальных температур для фотосинтеза (рис. 72) и его нижних температурных порогов, эти явления хорошо выражены у растений холодных местообитаний. Так, у альпийских и антарктических лишайников для фотосинтеза оптимальна температура около 5°С; заметный фотосинтез удается обнаружить у них даже при —10°С.
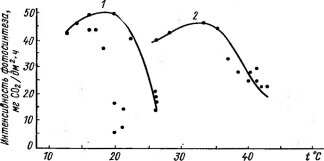
При сравнительно низких температурах лежит оптимум фотосинтеза у арктических растений, высокогорных видов, ранневесенних эфемероидов. Зимой при отрицательных температурах способны к фотосинтезу многие хвойные древесные породы. У одного и того же вида температурные оп- тимумы фотосинтеза связаны с изменением условий; так, у альпийских и арктических популяций травянистых многолетников — Оху- ria digyna, Thalictrum alpinum и других видов они более низкие, чем у равнинных. Показательно в этом отношении и сезонное смещение оптимума по мере повышения температуры от весны к лету (рис. 73, А) и снижения от лета к осени и зиме (рис. 73, Б).
При низких температурах для растений чрезвычайно важно сохранить достаточный уровень дыхания — энергетической основы роста и репарации возможных повреждений холодом. На примере ряда растений памирских высокогорий показано, что в этих условиях довольно интенсивное дыхание сохраняется после действия іемпературьі от —6 до —10°С (рис. 74).
Еще один пример устойчивости физиологических процессов к холоду— зимний и предвесенний подснежный рост у растений тундр, ¦высокогорий и других холодных местообитаний с коротким вегетационным периодом, обусловленным заблаговременной подготовкой. Это явление чрезвычайно ярко выражено у эфемероидов лесостепных дубовых лесов (пролески — Scilla sibirica, хохлатки — Gorydalis halleri, гусиного лука — Gagea lutea, чистяка — Ficaria
А
![]()
Б
![Сезонное смещение температурного оптимума фотосинтеза. А — полынь — Artemisia turanica (юго-западные Кызылкумы, по Захарьянцу И. Л. и др., 1973)]()
Рис. 73. Сезонное смещение температурного оптимума фотосинтеза. А — полынь — Artemisia turanica (юго-западные Кызылкумы, по Захарьянцу И. Л. и др., 1973):
/ — весна (конец апреля), 2 — лето;
Б — лавровишня — Laurocerasus officinalis (вечнозеленый кустарник, Австрия, по Pisek A. et al., 1969):
І — зима, 2 — лето
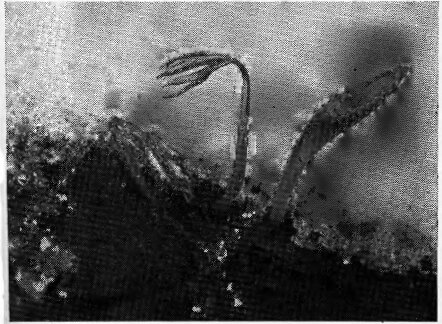
verna и др.), у которых уже в начале зимы начинается рост побегов со сформированными внутри бутонами (вначале в промерзшей почве, а затем над почвой, внутри снежного покрова — рис. 75). Не прекращается у них зимой и формирование генеративных органов. По мере приближения сроков снеготаяния скорость подснежного роста заметно возрастает. В пору раннего «предвесенья», когда лес кажется еще совсем безжизненным, под снеговым покровом над почвой уже возвышаются тысячи ростков пролески и гусиного лука,
-12 -10 -8 -6 -В -2 О
1 1 I 1 I L... і.
![Температурные зоны дыхания некоторых высокогорных растений Восточного Памира (по Семихатовой О. А., 1956). Зоны: / — нормального дыхания, II — обратимого изменения дыхания, III — подавленного]()
Рис. 74. Температурные зоны дыхания некоторых высокогорных растений Восточного Памира (по Семихатовой О. А., 1956). Зоны: / — нормального дыхания, II — обратимого изменения дыхания, III — подавленного
дыхания:
Виды: / — горец — Polygonum viviparum, 2 — первоцвет — Primula pamirica. З — полынь — Artemisia rhodantha, 4 — терескен — Eurotia ceratoides ![Подснежный рост ранневесенних эфемероидов в дубовом лесу Раскопка снега в марте. Видны ростки ветреницы лютичной и хохлатки Галлера (Ув.)]()
Рис. 75 Подснежный рост ранневесенних эфемероидов в дубовом лесу Раскопка снега в марте. Видны ростки ветреницы лютичной и хохлатки Галлера (Ув.)
достигающих к этому времени 2—7 см высоты и готовых начать цветение, как только сойдет снег. Образование хлорофилла у ранневесенних эфемероидов также начинается при низких температурах порядка 0°С, еще под снегом.
Экологические различия холодостойкости растений. В экологии и экологической физиологии в качестве одного из показателей устойчивости к холоду используется способность растения переносить низкую температуру в экспериментальных условиях в течение определенного срока. Накоплено много данных, позволяющих сравнивать растения различных по температурным условиям местообитаний. Однако эти данные не всегда строго сравнимы, поскольку температура, которую способно вынести растение, в числе прочих причин зависит и от продолжительности ее действия (так, небольшой холод порядка —3—5°С умеренно теплолюбивое растение способно вынести в течение нескольких часов, но та же температура может оказаться губительной, если будет действовать несколько суток). В большинстве экспериментальных работ принято охлаждение растений в течение суток или близкого срока.
Как видно из нижеследующих данных и рис. 76, холодостойкость растений весьма различна и зависит от. условий, в которых они обитают.
Холодостойкость растений' различных экологических групп (по Библю Р., 1965, с использованием данных разных авторов)
Уровень холодостойкости, °0
5—14
5—6
(в течение 2—4 суток), 15—17 (в течение 15—20 суток)
Группа растений
Водоросли тропических морей . . . Низшие термофильные грибы . . .
2—4 От 0 до 7
От —9 до—15
» — 5,5 » —13,5 »— 11,5 » —20
» _ 19 » _40
» — 2,5 » —5,5
» — 28 » —36
» — 34 » —47
Оранжерейные декоративные культуры (выходцы из тропических лесов)
Водные цветковые
Средиземноморские вечнозеленые деревья и кустарники
Лесные травы, зимующие подземные
органы
Лесные травы, зимнезеленые листья Лиственные деревья, зимующие почки
Лиственные деревья, распускающиеся листья
Карликовые кустарнички альпийской зоны
Один из крайних примеров холодостойкости — так называемый «криопланктон». Это снежные водоросли, живущие в поверхностных слоях снега и льда и при массовом размножении вызывающие его окрашивание («красный снег», «зеленый снег» и т. д.). В актив-
Хвойные деревья, верхняя граница леса х
ных фазах они развиваются при 0°С (летом на оттаявшей поверхности снега и льда). Пределы устойчивости к низким температурам от —36°С у Chlamydomonas nivalis до —40, —60°С у Pediast- rum boryanum, Hormidium flaccidum. Столь же велика холодостойкость фитопланктона полярных морей, нередко зимующего в корке льда.
Большой холодостойкостью отличаются альпийские карликовые кустарнички — Rhododendron ferrugineum, Erica сатеа и др. (—28,
0 -5
Тропики Субтропики
Области с мягкой зимой Области с суровой зимой
![Чувствительность к холоду древесных растений иа различных климатических областей (по Larcher W., 1971)]()
-10 -15 -20 -25 -30 -35 Г С
Рис. 76. Чувствительность к холоду древесных растений иа различных климатических областей (по Larcher W., 1971).
Степень повреждения в эксперименте: 1 — начало повреждения, 2 и 3 — гибель 50 и 100% экземпляров. Верхний край полосы соответствует наиболее чувствительному виду, НИЖНИЙ край — наименее чувствительному
—36°С), хвойные древесные породы: так, для сосны Pinus strobus в Тирольских Альпах в экспериментах отмечена рекордная температура: —78°С.
Совсем небольшая холодостойкость у растений тропических и субтропических областей, где они не испытывают действия низких температур (за исключением высокогорий). Так, для водорослей тропических морей (особенно мелководных районов) нижняя температурная граница лежит в пределах 5—14°С (вспомним, что для водорослей арктических морей верхняя граница составляет 16°С). Саженцы тропических древесных пород гибнут при 3—5°С. У многих тропических термофильных растений, например декоративных оранжерейных видов из родов Gloxinia, Coleus, Achimenes и др., понижение температуры до нескольких градусов выше нуля вызывает явления «простуды»: при отсутствии видимых повреждений через некоторое время останавливается рост, опадают листья, растения завядают, а затем и гибнут. Известно это явление и для теплолюбивых культурных растений (огурцов, томатов, фасоли).
Очень невелика устойчивость к холоду у термофильных плесневых грибов из родов Mucor, Thermoascus, Anixia и др. Они гибнут за три дня при температуре 5—6°С и даже температуру 15—17°С не могут выносить дольше 15—20 дней.
В связи с неравномерностью прихода солнечного тепла в разные сезоны года в умеренных и высоких широтах период, обеспеченный теплом для активной жизнедеятельности растений, ограничен лишь определенным отрезком года — вегетационным сезоном. В годовой динамике температур четко выражен более или менее продолжительный холодный период, когда преобладают отрицательные температуры и активная жизнедеятельность растений невозможна.
В качестве основной адаптации к этому сезонному ритму климатических условий у растений и других пойкилотермных организмов (а также некоторых гомеотермных) в годичном цикле развития чередуются активные и покоящиеся фазы. Однолетние растения после завершения вегетации переживают осень и зиму в виде семян, многолетние переходят в состояние осенне-зимнего покоя.
Состояние покоя у растений характеризуется рядом физиологических особенностей, дающих возможность переносить холод и все другие невзгоды осенне-зимнего периода. Потеря листьев при осеннем листопаде деревьев, кустарников, кустарничков, отмирание надземных частей многолетних травянистых растений — это сокращение транспирирующей поверхности, необходимое зимой, когда корни не способны пополнять большие потери воды, а также сокращение излишних расходов запасных веществ на дыхание листьев.
У зимующих частей растений уменьшается содержание воды (особенно ее свободной формы) и проницаемость цитоплазмы, резко замедляется скорость окислительно-восстановительных процессов (снижается интенсивность дыхания), значительно падает содержание ауксинов и других ростовых веществ. Все эти изменения в целом означают понижение интенсивности обмена веществ и блокирование (если не полное, то частичное) путей взаимодействия с окружающей средой, что уменьшает восприимчивость растений к неблагоприятным и повреждающим влияниям.
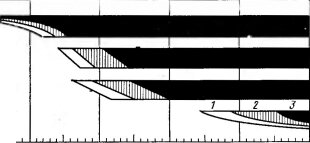
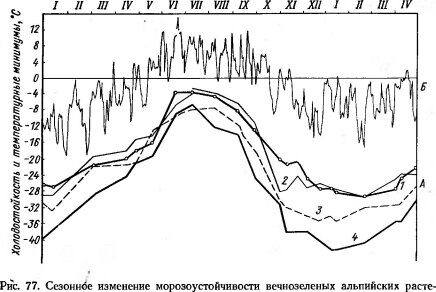
В состоянии покоя растения обладают повышенной устойчивостью к различным экстремальным факторам: нагреву, высушиванию и т. д., а также к ядам и другим неэкологическим воздействиям. Но особенно важно то, что в это время значительно увеличивается холодостойкость. Многочисленные исследования на самых разных растительных объектах позволили установить соответствие сезонного хода температур уровню холодостойкости (рис. 77). В состоянии покоя растения способны выдерживать длительное и сильное охлаждение. Не защищенные снеговым покровом почки лиственных деревьев и кустарников, хвоя и почки хвойных в областях с суровыми зимами выносят 40°-ные морозы. В опытах с глубоким охлаждением было показано, что ветви древесных пород в состоянии зимнего покоя выдерживают кратковременное пребывание при ¦—200°С, а покоящиеся семена и споры — от —190 до —197°С. По окончании покоя, когда в растении возобновляются активные процессы сокодвижения, деления клеток, роста и т. д., холодостойкость резко падает. Так, у европейских лиственных древесных по-
![]()
ний (А) и суточные минимумы температур в их местообитаниях (Б) (по Ulmer W.,
1937):
1 — Calluna vulgaris, 2 — Rhododendron ferruglneum, 3 — Loiseleuria procumbens, 4 — Pinus cembra ![Устойчивость к холоду зимующих почек ясеня Fraxinus ornus и ее сезонные изменения (по Mair В., 1968)]()
А Б Б Г
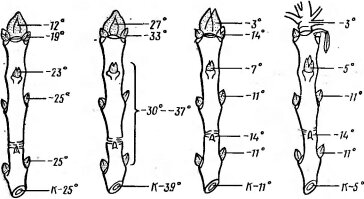
Рис 78. Устойчивость к холоду зимующих почек ясеня Fraxinus ornus и ее сезонные изменения (по Mair В., 1968). А — конец ноября; Б — декабрь; В — начало апреля (набухание); Г — середина апреля (развертывание). Цифрами с буквой К .обозначена устойчивость камбия
род зимующие почки могут вынести морозы от —20 до —30°С, а для распускающихся листьев уже —3, —4СС могут быть гибельными (рис. 78). Известно, как легко побиваются поздневесенними заморозками почки, тронувшиеся в рост.
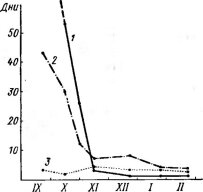
![Динамика глубины осенне- зимнего покоя у дубравных растений: I — Scilla sibirica, 2 — Asarum europaeum]()
Месяцы
Рис. 79. Динамика глубины осенне- зимнего покоя у дубравных растений: I — Scilla sibirica, 2 — Asarum europaeum,
З — Aegcpodium podagrarla. По оси ординат — число дней от перенесения из природных условий в тепло до начала развития
В течение всего осенне-зимнего периода покоящиеся растения в большинстве случаев не обнаруживают никаких внешних признаков изменения. Однако в этот период в растениях происходят глубокие внутренние сдвиги, и их состояние в конце зимы сильно отличается от осеннего. Это хорошо видно по изменению способности растений реагировать на оптимальные тепловые условия (рис. 79). Если на первом этапе покоя (осенью или в начале зимы) поставить в воду срезанную ветку дерева или выкопать из-под снега травянистый многолетник и поместить их в тепло, то почки не раскрываются очень долго (а у некоторых не раскрываются совсем). В это время не удается вывести растения из состояния покоя и другими воздействиями, например теплыми ваннами, химическими или механическими раздражителями. Эта фаза получила название глубокого, или органического, покоя. Уже в декабре — начале января срок от момента внесения в тепло до распускания почек сокращается, т. е. покой становится менее глубоким. Начиная с середины — конца января, растения реагируют на действие тепла очень быстро: почки распускаются через 2—3 дня, и в последующие месяцы это состояние уже не меняется. Иными словами, растения готовы к возобновлению развития, глубокий покой окончен, но начало развития в природе невозможно из-за низких зимних температур, и потому эту фазу называют вынужденным покоем.
Сопоставление сроков смены глубокого и вынужденного покоя с ходом природных температур показывает, что у большинства растений умеренных широт покой, по существу, кончается не весной, а в самый разгар зимы, тогда, когда обычно наступают сильные холода. Однако не следует забывать об особенностях того природного режима температур, на фоне которого складывалась смена фаз покоя. Осенний период с его неустойчивым температурным режимом, частой сменой похолоданий и потеплений опасен для растений тем, что кратковременные потепления могут вызвать возобновление роста и развития и стать толчком к началу новой вегетации. Нетрудно представить гибельные последствия таких про-
вокаций в последующие зимние месяцы. Адаптивный смысл глубокого покоя как раз и заключается в защите против возможных осенних провокаций преждевременного начала развития. Когда наступают устойчивые холода, необходимость в глубоком покое отпадает, поскольку теперь от возобновления развития растения достаточно надежно предохраняют низкие зимние температуры. Не случайно у большинства растений умеренных широт переход от глубокого покоя к вынужденному обычно совпадает с тем временем, когда прекращаются оттепели и устанавливаются устойчивые низкие температуры.
Фаза глубокого покоя выражена не у всех растений: некоторые травянистые многолетники способны продолжать развитие в тепле и осенью (рис. 79, 3), т. е. в течение всего осенне-зимнего периода в природных условиях они находятся в состоянии вынужденного покоя. Обычно это сравнительно теплолюбивые растения, и от осеннего возобновления вегетации их предохраняют высокие температурные пороги начала развития.
Подготовка растений к состоянию покоя происходит заблаговременно (так же, как и у животных, впадающих в спячку или диапаузу). Задолго до наступления холодов начинаются пожелтение и сбрасывание листвы у деревьев и кустарников, процессы «вызревания» побегов (одревеснение, утолщение пробкового слоя). У многолетних травянистых растений и озимых культур происходит закаливание— накопление растворимых углеводов и других защитных веществ, повышающих морозостойкость. Зимующие почки водных растений опускаются на дно водоемов. Эта заблаговременность обеспечивается природным сигналом, вслед за которым неизбежно наступает понижение температуры, — изменением фотопериода, т. е. осенним сокращением дня. Экспериментально показано, что именно осенний фотопериод вызывает начало листопада, увеличение морозостойкости (рис. 80), вызревание древесных побегов и другие формы подготовки растений к покою, так же как и предзимние явления в животном мире: отлет птиц, впадение насекомых в диапаузу, животных в спячку.
Сокращение дня служит очень удачным сигналом, поскольку оно ежегодно наступает с астрономической точностью. Однако это не означает, что и осенние явления в растительном мире каждый год наступают в одни и те же календарные даты. Это связано с тем, что приуроченность осенних явлений к конкретным условиям года регулируется не только световым, но и температурным фактором, который вносит определенную «поправку» к восприятию фотопериода. Есть данные, показывающие, что одна и та же длина дня в теплую осень воспринимается как более длинная (т. е. как продолжение лета), а при похолоданиях — как более короткая.
Окончание состояния покоя и начало нового вегетационного периода (начало сокодвижения у деревьев, раскрывание почек, отрастание побегов, бутонизация и т. д.) происходят под действием весеннего повышения температуры, т. е. гораздо позже, чем заканчивается фаза глубокого покоя у растений (в середине зимы).
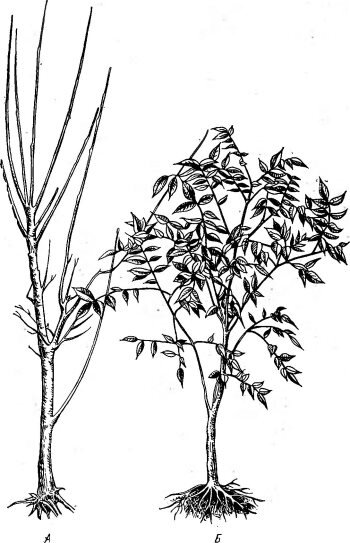
![Пример влияния фотопериодического режима на подготовку растений к зиме (по Мошкову Б. С., 1Э61)]()
Рис. 80. Пример влияния фотопериодического режима на подготовку растений к зиме (по Мошкову Б. С., 1Э61). Саженцы амурского бархата в Ленинграде, выращенные на естественном дне (А), полностью вымерзли при перезимовке; выращенные на искусственно укороченном дне (Б) сохранились
Что же здесь служит сигнальным фактором? Им могло бы быть предвесеннее удлинение фотопериода, однако большинство растений в это время находится в безлистном состоянии, а фотопериоди- ческая информация, как известно, воспринимается зеленым листом. Понять, какие влияния среды необходимы для выхода растений из глубокого покоя, помогает анализ поведения растений при различном темпера- «.
Рис. 81. Опыт, демонстрирующий необходимость осеннего холода для прохождения годичного цикла развития (на примере хохлатки Corydalls halleri).
А — растения, содержавшиеся осенью при температуре 5°С, нормально цветут и плодоносят при выгонке в тепле; Б — задержка развития после теплой осени (18°С).
Фенофазы: / — прорастание, 2 — бутонизация, 3 —
цветение и плодоношение, 4“ отмирание. Жирная линия — период холода
18°С
el
5'С
16°С
II III Месяцы
IX X XI XII
турном режиме в экспери- менте (рис. 81). Если, начиная с осени, содержать растения не на естественном температурном фоне, а при комнатной температуре (иными словами, лишить его холода осенью и в начале зимы), то выход из состояния покоя сильно задерживается и дальнейшее развитие тормозится (в частности, у многих видов выпадают генеративные фазы). Потребность растений умеренных широт в осенних пониженных температурах для нормального цикла
развития была многократно показана на самых разных объектах Е Для некоторых культурных растений (например, озимых злаков) она получила название яровизации: если проросшие семена не подвергнутся действию осеннего холода, в следующее лето не наступит колошение.
В экспериментах показано, что потребность в пониженных температурах у зимующих растений приурочена имени» к осенним месяцам. Продолжительность действия холода должна быть не менее 2—3 месяцев, а температура не выше 3—5°С.
В природных условиях необходимую дозу холода растение получает к началу зимы и, возможно, это является своеобразным сигналом окончания того периода, когда необходимо защитное действие глубокого покоя против преждевременных потеплений, провоцирующих развитие. По-видимому, здесь действует механизм накопления информации из внешней среды до определенной величины, после чего эта информация (доза холода) срабатывает как сигнал для перехода из состояния глубокого покоя в состояние вынужденного.
Потребность в пониженных температурах в осенний период и в
1 Аналогичное явление, получившее название «реактивации», существует и у других живых организмов — насекомых, впадающих в осенне-зимнее покоящееся состояние (диапаузу).
тепле в период вегетации означает, что для растений умеренных широт имеет значение не только общий уровень тепла, но и определенное распределение его во времени. Так, если в опыте (рис. 82) растениям дать одинаковое количество тепла, но по-разному распределенное: одному теплое лето и холодную зиму, а другому соответ-
![Схема сезонного термопериодизма на примере пролески Scilla sibirica]()
Рис. 82. Схема сезонного термопериодизма на примере пролески Scilla sibirica. А — нормальная сезонная смена температур: теплое лето и холодная осень;
Б — постоянная средняя температура.
Фенофазы: / — прорастание, 2 — бутонизация, 3 — цветение н плодоношение, 4 — отмирание. Жирные линнн — периоды холода или средней температуры. Выгонка
при 18°С
ствующую постоянную среднюю температуру, то нормальное раз- витие будет только в первом случае, хотя общая сумма тепла в обоих вариантах одинакова. Потребность растений умеренных широт в чередовании в течение года холодных и теплых периодов получила название сезонного термопериодизма.
Кроме того, для растений небезразлично и распределение температур в течение суток: известно* например, что развитие многих культурных растений более успешно идет не на постоянном температурном фоне в течение вегетации, а при ночных понижениях температуры. Это явление, получившее название суточного термопериодизма, впервые было обнаружено в Нидерландах на примере культуры томатов в закрытом грунте; лучший урожай получали более беспечные хозяева, не топившие свои теплицы по ночам.