ВЛИЯНИЕ ЗАСОЛЕНИЯ НА РАСТЕНИЯ
В той или иной мере засолено около 25% всех почв нашей планеты. Избыток солей в почвенном растворе токсичен для большинства растений. Наиболее вредны легкорастворимые соли, без труда проникающие в цитоплазму: NaCl, MgCl2, СаС12.
Менеетоксичны труднорастворимые соли: CaSO}, MgS04, СаСОз.
Как показано в многочисленных экспериментах, особенно сильно действует на растения хлоридное засоление, в то время как сульфатное менее вредно. Меньшая токсичность сульфатного засоления, в частности, связана с тем, что в отличие от иона С1~ ион S04_ в небольших количествах необходим для нормального минерального питания растений, и вреден только его избыток.
Избыточная концентрация солей оказывает как осмотическое действие, нарушающее нормальное водоснабжение растений, так и токсическое, вызывая отравления. В частности, отравление возникает в результате резкого нарушения азотного обмена и накопления продуктов распада белков. Сильное засоление замедляет синтез белков, подавляет процессы роста. Засоление почвы угнетающе действует и на почвенные микроорганизмы (в том числе на те группы, жизнедеятельность которых весьма существенна для высших растений). В жарком и сухом климате с преобладанием в почве восходящего тока воды засоление встречается очень часто, на огромных площадях (большая часть территорий степной, полупустынной и пустынной зон). Здесь источником засоления служит не только приток солей из грунтовых вод, но даже атмосферные осадки, несущие ничтожные количества минеральных солей. Засоление в таких местностях может быть вызвано и неумелым орошением.
Во влажном климате при постоянном промывании почв накопления солей не происходит, и их содержание в почвенном растворе не превышает сотых или тысячных долей процента. Исключение составляют лишь почвы, постоянно увлажняемые сильно засоленными выходами грунтовых вед или морскими приливами (по побережьям Балтийского, Белого и других северных морей).
Еще один источник засоления — отложение морских солей ветром на приморских скалах (импульверизация), в большой степени влияющее на состав и размещение прибрежной растительности.Среди разных типов засоленных почв .основные — солончаки и солонцы, имеющие неодинаковый солевой и водный режимы.
Солончаки — почвы, постоянно и сильно увлажненные солеными водами вплоть до поверхности («мокрые» солончаки, «соленые грязи»), например вокруг горько-соленых озер. Летом с поверхности солончаки высыхают, покрываясь корочкой солей. Концентрация солей в почвенном растворе достигает нескольких десятков процентов; ионы натрия находятся не только в растворе, но и насыщают коллоиды почвенного поглощающего комплекса.
Солонцы с поверхности не засолены, верхний слой выщелоченный, бесструктурный. Нижние горизонты уплотнены и насыщены ионами натрия, при высыхании растрескиваются на столбы, глыбы и т. д. Водный режим характеризуется резкими изменениями (весной вследствие водонепроницаемости нередко наблюдается поверхностное застаивание влаги, летом — сильное пересыхание). Есть ряд промежуточных типов почв: солончаковатые солонцы, солончаковатые почвы, солонцеватые и т. д. Часто они располагаются вперемешку, небольшими участками, создавая солончаково-солонцовые комплексы (Прикаспий, Казахстан и т. д.).
Ка засоленных почвах поселяются растения, приспособившиеся к высокому содержанию солей,— галофиты. Они отличаются специфическим видовым составом: так, есть целые «галофильные» семейства (маревых — Chenopodiaceae, свинчатковых — Plumbagi- пасеае, франкениевых — Frankeniaceae, тамариксовых — Tamarica- сеае). Флора галофитов особенно богата и разнообразна в аридных зонах. В отличие от галофитов растения незаселенных местообитаний называют гликофитами.
В зависимости от морфофизиологических особенностей и путей адаптации к засолению различают несколько групп галофитов (по Генке- лю П. А.):
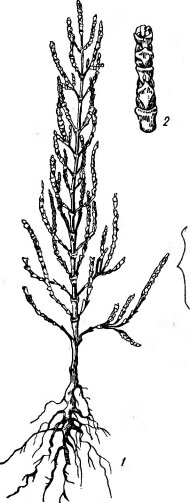
- Эугалофиты (солянки). В основном это обитатели солончаков или солончаковых участков засоленных комплексов. Примеры — солерос европейский (Salicornia еи- гораеа), сарсазан шишковатый (На- locnemum strobilaceum), виды родов Salsola, Petrosimonia, Suaeda, Obio- ne и др. Характерен внешний облик этих растений. Некоторые из них имеют суккулентные черты (рис.
- : редуцированные листья, мясистые членистые стебли, по периферии которых располагается ассимиляционная ткань—двухслойная палисадная паренхима, а центральная часть . занята сочной водозапасающей j тканью. У других (Obione, Suaeda) (.листья обычные, пластинчатые, но с ^чертами «галоморфной» структуры: ¦сильно утолщенной и несколько сук- кулентной листовой пластинкой, сравнительно крупными клетками.
Связь этих черт с засоленностью почв была показана еще в конце прошлого века в опытах, в которых резкие анатомо-морфологические изменения растений вызывались поливом растворами сернокислых и хлористых солей. В более поздних работах (в том числе на культурных растениях) также показано, что при из- 1gt;ис. 139. Однолетний галофит— бытке солей наблюдается утолщение солерос Salicornia еигораеа: листовой пластинки за счет разрас-
/ — целое растение, 1 — отдельная не- „
точка (увеличено) тания палисадной паренхимы, сни-
жается число устьиц: лист становится более суккулентным, а размер листовой пластинки уменьшается. Такие явления наблюдаются при разных типах засоления (рис. 140).
В связи с высокой солеустойчивостью цитоплазмы эугалофиты способны поглощать и накапливать большие количества солей (до 45—50% от массы золы). По современным представлениям, соли
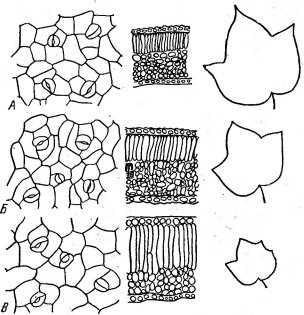
Рис. 140. Изменение строения листа хлопчатника при засолении (по Строгонову Б. П., 1973).
А — контроль; Б — сульфатное и В — хлоридное засоление
накапливаются не только в цитоплазме, но и в «инертных» образованиях клетки, которые используются как вместилища для балластных солей.
Чем засоленнее почва, тем больше солей содержат н растения-соленакопители. Состав ионов в теле растения обычно соответствует их соотношению в почве, хотя есть примеры избирательного накопления: так, злаки и осоковые накапливают больше К, чем Na, маревые — Na, Cl и органические анионы (главным образом, оксалат), крестоцветные — Na и S04. Солерос концентрирует в тканях значительное количество солей даже при малом содержании их в почве. Некоторые галофиты-соленакопители при массовом произрастании раньше даже служили источником сырья для получения соды и поташа (что отразилось и в их латинских названиях — Salsola kali, S. soda, род Kalidium). Опад эугалофи- тов способствует засолению поверхностного слоя почвы: хорошо известно образование сцементированной солями корки под кронами саксаулов в Средней Азии, а в австралийских полупустынях существенно засоляют почву виды рода Atriplex. Под зарослями галофитов происходит также подщелачивание почвы.- Криногалофиты (солевыделители). Эти растения способны выделять наружу избыток соли в виде солевого раствора через особые железки на листьях. Таковы тамариксы, франкения, многие кермеки (Limonium). (Ботаники хорошо знают, как при сборах гербария трудно высушить ветку тамарикса с листьями, покрытыми солью и потому «отмокающими» в бумаге.) Сухой порошок соли сдувается с листьев ветром, осыпается или смывается дождем. По строению листа многие криногалофиты близки к мезофитам.
- Гликогалофиты. Сюда относятся растения ксерофиль- ного облика (например, многие полыни, покрывающие огромные территории в степной и полупустынной областях). Корневая система гликогалофитов малопроницаема для солей, поэтому даже на сильно засоленных почвах в тканях растений соли не накапливаются. Механизм этого «корневого барьера» еще не ясен: одни авторы придают значение утолщению стенок клеток корня, другие— биохимическим процессам, обеспечивающим задержку вред ных солей, третьи — способности растений существовать при очень экономном водном режиме и потому меньше поглощать «балластные» ионы.
Следует еще упомянуть о группе растений, избегающих засоления благодаря глубокой корневой системе: хотя они и могут расти на сильно засоленных почвах, их сосущие корни располагаются в глубоких малозасоленных горизонтах.
Таков, например, тростник (Phragmiies communis). Химические анализы тростни ка, растущего в пойме Сырдарьи на почвах, очень различных по составу и степени засоления, показывают большое однообразие состава надземных органов и особенно корней, в то же время состав растущих здесь же однолетних солянок с неглубокими корнями сильно зависит от свойств верхних засоленных горизонтов. Аналогичны тростнику кендырь сибирский (Аросупит sibiricum) и другие виды. Эту группу иногда называют псевдогалофи- т а м и.Галофиты имеют ряд биохимических особенностей, способствующих «обезвреживанию» вредных солей и выведению их из основного обмена. Это обеспечивается избирательным связыванием ионов белками цитоплазмы и органическими кислотами, а также накоплением веществ, выполняющих защитную функцию (некоторые углеводы, аминокислоты, пигменты и др.).
У многих галофитов проницаемость цитоплазмы для солей заметно снижена, она более гидрофильна, содержит много коллоид но-связанной воды, что способствует экономному расходу воды и меньшему поглощению солей.
Концентрация солен в клеточном соке галофитов обусловливает высокое осмотическое давление порядка 5000 кПа и более (в то время как у других растений с высоким осмотическим давлением— ксерофитов — оно достигается благодаря сахарам). Повышение осмотического давления обеспечивает возможность всасывания корнями воды из концентрированного почвенного раствора.
Следует подчеркнуть, что для многих галофитов повышенная концентрация солей в почве — необходимый и благоприятный фактор. Так, опыты культуры солероса и морского подорожника (Plantago salsa) на разных почвенных растворах показали, что для их роста оптимальна концентрация поваренной соли 2—3%, в то время как при выращивании этих растений в пресной воде наблюдалось угнетение. Такой же «положительный солевой эффект» отмечен для сарсазана, лебеды и др. Он проявляется как у взрослых растений, так и у прорастающих семян.
Экспериментально показано благоприятное влияние умеренного засоления на разные стороны физиологии галофитов (что особенно заметно при сравнении с гликофитами).
Например, при выращивании галофитов на питательной смеси с добавлением NaCl активность ферментов, ответственных за солевой обмен, остается высокой даже при концентрациях порядка 12 г/л (а у гликофитов падает уже при 3 г/л), усиливается азотный обмен и синтез белков. Происходят сдвиги и в почвенном питании: так, поглощение меченого фосфора у галофитов максимально при pH 8—9 и стимулируется ионами натрия, а у гликофитов для этого процесса оптимальна область pH 5—6, натрий же действует на него угнетающе.Устойчивость галофитов к засолению демонстрируется также следующим опытом: у их листьев, погруженных черешками в солевой раствор, сохраняется нормальный фотосинтез, в то время как у листьев гликофитов обнаруживается угнетение фотосинтеза, разрушение хлорофилла, «солевые ожоги» (отмирание части листовых пластинок).
Однако и у самых солеустойчивых галофитов выносливость к засолению имеет предел: при резком избытке солей они испытывают угнетение. Это хорошо показано на примере солероса и других солянок в условиях очень засоленных почв.
Произрастая на сравнительно влажных солончаках, многие эугалофиты недостаточно выносливы к засухе. На более сухих почвах (например, сильно пересыхающих летом солончаковатых солонцах) поселяются галофиты с теми или иными ксероморфны- ми чертами: кокпек — Atriplex сапа и лебеда бородавчатая —
к. verrucifera с опушенными листьями и глубокой корневой системой, биюргун — Anabasis salsa с утолщенным двухслойным эпидермисом и др. На сухих столбчатых солонцах преобладают виды с ярко выраженными ксероморфными чертами: сильно опушенными и рассеченными листьями (полыни — малоцветковая и Лерха — Artemisia pauciflora, A. lercheana, ромашник — Pyrethrum ach.il- leifolium) или с очень мелкими игловидными листьями (Campho- rosma monspeliacum). На этих же почвах способны расти и некоторые выносящие засоление степные ксерофиты, например
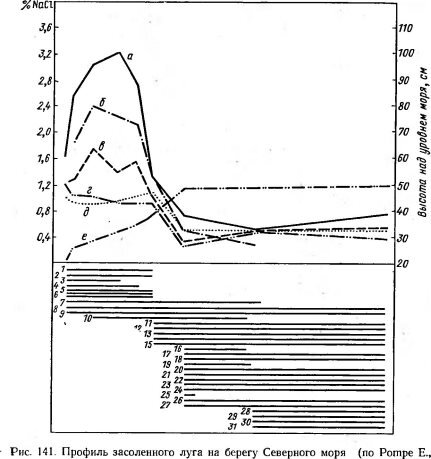
1940).
Содержание NaCl в горизонтах почвы: а—1—2 см, 6 — 6 см, в—15 см, г — 30 см, д 45 см, е — положение точек профиля над уровнем моря.
Виды: / — водоросли, 2 — Atropis distans, 3 — Spergularia satina, 4 — Salicornia herbacea,
5 — Glaux maritima, 6 — Aster tripolium, 7 — Triglochin maritima, 8 — Juncus gerardii,
9 — Plantago maritima, 10 — Armeria vulgaris var. maritima, 11— Plantago coronopus, 12~~Trifolium fragiferum, ІЗ — Festuca arundinacea var. baltica, 14 — Festuca rubra,
/*» — Agrostis alba, 16 — Apium graveolens, 17 — Leontodon autumnalis, 18 — Potentilla anserina, 19 — Potentilla reptans, 20 — Lotus tenuifolius, 21 — Erythraea pulchella, 22 — Sa- gina nodosa, 23 — Sieglingia decumbens, 24— Trifolium repens, 25 — Beilis perennis, 26 — Odontites litoralis, 27 — Taraxacum officinale var. palustre, 28 Carex goodenoughii,
29 — Carex distans, 30 — Bupleurum tenuissimum, 31 — Brunella vulgaris
типчак — Festuca sulcata. Можно, таким образом, проследить ряд переходов от настоящих галофитов к галоксерофитам и ксерофитам.
Есть и другая промежуточная группа — галомезофиты Это в основном растения лугов, произрастающие на слабозасолен- ных почвах: в поймах лесостепных и степных рек, в местах с подтоком засоленных грунтовых вод. По внешнему облику и строению они не отличаются от обычных луговых мезофитов. Таковы лисохвост вздутый — Alopecurus ventricosus, бескильница расставленная— Puccinellia distans, клубнекамыш морской — Bolboschoenus maritimus, морковник Бессера — Silaus besseri и др. Некоторые из них имеют повышенное содержание солей (что придает сену с таких лугов солоноватый привкус, привлекательный для животных) и способны выделять их избыток в процессе гуттации.
Своеобразную группу составляют приморские галофиты и га- ломезофиты — растения «маршей» (приморских лугов, рис. 141), литоралей, дюн и зон, контактных с дюнами, приморских скал. В этих местообитаниях избыток солей, поступающий в почву с. морскими приливами, из засоленных грунтовых вод или с ветром, для одних видов сочетается с избытком увлажнения (и даже временным анаэробиозом при заливании), для других, растущих на повышенных и песчаных субстратах,— с ограниченным количеством доступной влаги. Видовой состав приморской флоры (в пределах Европы) довольно однообразен. Здесь можно найти и типичный галофит — солерос — Salicornia еигораеа, и виды с гало- морфными чертами — утолщенными суккулентными листьями (Honkenya peploides, Glaux maritima, Mertensia maritima и др.). и растущие на засоленных песках галопсаммофиты с теми или иными ксерофильными чертами (достаточно вспомнить встречающиеся на побережье Финского залива голубые заросли крупного злака волоснеца гигантского — Elymus giganteus с листьями, покрытыми сизым восковым налетом). Как показали исследования приморских растений на побережье Литвы, они имеют повышенную концентрацию клеточного сока, высокое осмотическое давление (от 2200 до 5300 кПа) и накапливают в надземных частях довольно значительные количества минеральных солей (12—22% от массы золы). Иногда вместе с приморскими галофитами растут и гликофиты, обладающие повышенной выносливостью к засолению: например, обычный вид, обитающий на побережьях пресноводных водоемов, частуха — Alisma plantago-aquatica, выносит pH почвы от 2,0 до 12,0 и концентрацию солей до 3%.
Особую группу среди растений засоленных местообитаний составляют мангровые заросли, или мангры,— затопляемые леса тропических побережий в зоне морских приливов или в эстуариях тро1 пических рек. Сильное засоление в этих местообитаниях создается благодаря действию морской воды, накоплению солей на берегах, приносу солевой пыли ветрами. Из древесных пород в манграх наиболее распространены роды Avicennia, Rhizophora и др.
Адаптации к засолению проявляются у них в некоторых физиологических механизмах, обеспечивающих избирательное поглощение ионов: например, более токсичный натрий поглощается меньше, чем калий. Избыток солей удаляется путем их активного выделения через особые устьица, поэтому нижний эпидермис листьев часто покрыт солевой корочкой. В том, что эти соли действительно выделены листом, а не осаждаются на его поверхности извне, убеждает быстрое восстановление солевой корочки после промывки интактных (не отделенных от растения) листьев, в том числе в «ветровой тени», где осаждение солевой пыли исключено. На примере Avicennia, растущей на побережьях Восточной Африки, показано, что выделяемый листом раствор содержит в основном ионы хлора, натрия, калия, причем в этом растворе их концентрация выше, чем в океанической воде. Высказываются предположения о том, что мангровые деревья способны поглощать из океанического «раствора» пресную воду путем простой (неметаболической) фильтрации. Осмотическое давление у мангровых древесных пород высокое, порядка 5000—7000 кПа. Возможны его колебания: так, на о. Ява у Avicennia в конце отлива осмотическое давление было 16 300 кПа, а в конце высокого прилива — 8200 кПа. Показано, что фотосинтез у мангровых деревьев идет вполне нормально при высоких концентрациях солей, которые у гликофитов сильно его подавляют.
Существует еще одна разновидность местообитаний, где необходима адаптация растений' к солености,— это воды морей и океанов. Напомним, что воды Мирового океана, содержащие 3—4% солей, составляют около 97% воды нашей планеты. Однако адаптации морских растений к засолению изучены еще очень мало.
Некоторые данные свидетельствуют о том, что морские водоросли способны накапливать в тканях большие количества солей (до 60% от сухой массы). Многие виды обнаруживают признаки угнетения при выращивании их в опресненной среде; так, для ряда галофильных водорослей оптимальна концентрация раствора NaCl в пределах 3—12%, в пресной же воде у них нарушается жизнедеятельность или они гибнут. В этом смысле некоторые авторы считают морские растения облигатными галофитами, а наземные— только факультативными, поскольку они способны жить и на незасоленных почвах.
Интересно, что анатомо-физиологические изменения в зависимости от концентрации солей у водных и наземных растений в известной степени сходны: так, у пресноводных водорослей при переносе в растворы солей увеличиваются размеры клеток, а также размеры ядра и ядрышка (как у галофильных суккулентов). Морские водоросли, выращенные в растворах более высокой, чем морская вода, концентрации, отличаются от типичных форм примерно теми же чертами, что и галофильные цветковые растения от обычных: более рыхлым строением тканей, меньшим развитием механических элементов, меньшим разветвлением.
В последние десятилетия возникла новая проблема, связанная с солеустойчивостью растений,— накопление солей в почвах вдоль дорог и улиц (в результате использования соли для удаления снега). Так, зимой 1968 г. в северных штатах США на шоссейных дорогах было использовано 6 млн. т соли (95% NaCl + 5% КС1). В районе Новой Англии за зиму на дорогах остается около 6 кг соли на погонный метр каждой стороны. Отсюда начинается засоление соседних территорий, в связи с чем в придорожных посадках необходим подбор солеустойчивых пород. В США от солей особенно страдают клен сахарный, тсуга канадская, сосны (угнетение роста, пожелтение, гибель). Неожиданно устойчивыми оказались яблони (Malus ргаесох) и цитрусовые; травы более солеустойчивы, чем деревья.
Засоленные почвы улучшают в основном специальной мелиорацией (промывкой и дренажем, химическим удалением поглощенного натрия). Возможно и применение растений, так называемая фитомелиорация, вследствие которой уровень засоленных грунтовых вод опускается, и почва рассоляется.
псаммофиты и ЛИТОФИТЫ
В морфологии и биологии псаммофитов и литофитов сказывается прежде всего своеобразие механического состава почв и обусловленная им экологическая обстановка.
Особенности песка как почвенного субстрата и растений сыпучих песков — псаммофитов. Хотя представление о песках связывается в основном с пустынями, песчаные массивы распространены не только в аридных, но и в более умеренных зонах. По происхождению пески могут быть приморскими, приозерными, приречными (и потому приурочены к берегам этих водоемов); связанными с деятельностью ледниковых потоков (современных или древних)—флювиогляциальными, или с деятельностью ветра— эоловыми. Различают сыпучие, или подвижные, пески, свободно перевеваемые ветром (например, подвижные пустынные барханы или подвижные дюны на морских побережьях) и пески, закрепленные растительностью.
Песок как субстрат имеет ряд особенностей, вызывающих необходимость адаптации со стороны растений. Во-первых, это тепловой режим: обладая малой теплоемкостью и высокой теплопроводностью, песок способен к сильному нагреванию и резким суточным колебаниям температуры (особенно на поверхности). В пустынях Средней Азии в жаркие дни температура поверхности песка доходит до 70—78°С при температуре воздуха в тени 40—43°С (на высоте 2 м). Отсюда и сильное нагревание приземного слоя воздуха.
Водный режим растений на песках складывается не совсем благоприятно. Это объясняется тем, что осадки быстро и беспрепятственно просачиваются вниз, так как водоудерживающая сила песка очень низкая, а скважность и водопроницаемость очень высокие. В связи с ничтожной капиллярностью песка обратный восходящий ток от грунтовых вод чрезвычайно мал. Эти особенности песчаных почв приводят к тому, что в поверхностных горизонтах для корневых систем растений создаются условия весьма скудного водоснабжения
В летние месяцы в среднеазиатских пустынях верхние слои песка иссушены почти полностью. К концу вегетационного сезона иссушение доходит до глубины 50—60 см. Вместе с тем на песчаных почвах создаются благоприятные условия для накопления влаги атмосферных осадков в глубоких горизонтах. Следует также отметить, что пески имеют очень небольшую величину мертвого запаса влаги (коэффициент завядания): например, для подвижных, хорошо перевеянных песков она составляет всего около 1%. При небольшом количестве осадков, выпадающих в пустынных областях, верхние слои песка промачиваются дождями неглубоко (до 1 м), поэтому гравитационная вода не доходит до уровня грунтовых вод, а образует «подвешенный» влажный горизонт, за счет которого и существует большая часть растительности пустынь. Для кустарников с глубокой корневой системой существенным источником водного питания может служить капиллярная кайма над грунтовыми водами. Иногда (особенно в осенние месяцы) растения могут дополнительно использовать влагу, которая образуется в результате внутрипочвенной конденсации.
Поскольку пески всегда хорошо промыты и поглощающий комплекс их невелик, содержание солей здесь очень мало. Количество органических вешеств в песках также невелико, особенно в пустынях со скудной растительностью, где поступление в почву органических остатков ограничено. Особенно бедны ими незакрепленные барханные пески, лишь под кронами редких пустынных кустарников и деревьев несколько повышено содержание гумуса, соединений азота, минеральных солей.
Наиболее яркая экологическая особенность песка как субстрата для растений — его подвижность, обусловленная весьма слабой связью между почвенными частицами.
В сухом состоянии перевеянные пески рыхлы, сыпучи и совершенно бесструктурны. Примеры последствий подвижности песка хорошо известны из истории среднеазиатских цивилизаций (погребение целых городов, пахотных массивов и т. д). В песчаных пустынях или на приморских дюнах непрерывный перенос песка ветром — фактор не эпизодический, а постоянный, и без приспособления к нему растения не могут существовать. Постоянная угроза для пустынных растений (не только мелких травянистых, но и крупных деревьев и кустарников) — частичное или полное погребение их под толщей песка (рис. 142). Существует и противоположная опасность — выдувание и оголение корней, грозящее им и всему растению засыханием (рис. 143). В качестве защиты у пустынных псаммофитов выработалась способность при погребении образовывать придаточные корни от ствола на любой высоте. Так, кустарники из рода джузгун (Calligonum) при засыпании развивают сильные придаточные корни на стеблях выше корневой шейки, а быстрый рост стеблей дает им возможность обогнать наступающие барханы. Часто от большого куста 3— 4-метровой высоты на поверхности бархана остается только щетка зеленых побегов, но благодаря придаточным корням куст продолжает жить. При отступлении песка придаточные корни обнажаются и остаются свидетелями бывшего уровня погребения — иногда в середине кроны или даже близ ее вершины (рис. 143). У пустынных злаков из рода Aristida (A. karelini, A. pennata) при раскопках видно несколько ярусов корней, располагающихся друг
Рис. 142. Засыпание пустынного кустарника движущимся барханом (по Петрову М. П, 1950). Видно образование придаточных корней

Рис. 143. Засохшие придаточные корни, сохранившиеся в кроне песчаной акации после отступания бархана
над другом на расстоянии 3—10 см, — свидетельство неоднократного засыпания, при котором корни развиваются на новой высоте.
Приспособление к возможному обнажению корней — образование на корнях защитных футляров из пробковой ткани или из песчинок, сцементированных выделениями корней (рис. 144). Такие корневые чехлики встречаются не только у настоящих пустынных
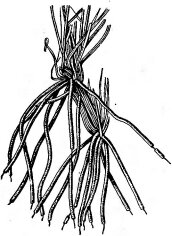
Рис. 144. Корни пустынного злака— Aristida karelini (по Петрову М. П., 1950). Видны корневые чехлики из сцементированных песчинок
растений, но и у видов, населяющих в наших широтах приречные и приморские пески, например у овсяницы Беккера (Festuca beckeri).
У многих псаммофитов подземные части имеют приспособления, направленные на то, чтобы как можно прочнее закрепиться в подвижном субстрате: распространены длинные, сильно разветвленные корневые системы экстенсивного типа (рис. 145, 146). Часто они расположены почти горизонтально в поверхностных слоях песка (например, корни у видов рода Aristida, см. рис. 146, длинные шнуровидные корневища осоки вздутой — Carex physodes; из более северных обитателей песков такие корневища имеет вейник наземный— Calamagrostis epigeios и др.).
Большую опасность для растений подвижных песков представляет погребение плодов и семян. В связи с этим у многих видов семена имеют приспособления к переносу ветром («парусные» выросты у солянок и саксаулов, перистые ости у се- лина, вздутия у осоки песчаной). Весьма специфическое «псаммо- фильное» приспособление—-развитие упругих ветвистых щетинок на семенах джузгунов (один из них получил название «голова Медузы», см. рис. 130); они облегчают прыгучесть семян и передвижение их ветром вместе с массами песка. Семена пустынных псаммофитов устойчивы к высокой температуре и долго сохраняют жизнеспособность даже при засыпании песком.
В связи с большими затруднениями в водоснабжении (особенно в областях пустынь) у псаммофитов выработались черты ярко выраженной ксероморфной организации. Среди них много суккулентов (именно в песчаных пустынях разнообразие крупных суккулентов наиболее велико). Большинство древесных и кустарниковых пород песчаных пустынь безлистны (саксаулы, джузгуны, солянки, биюргуны); фотосинтез у них осуществляется зелеными веточками цилиндрической формы. У других листья сильно редуцированы (песчаная акация). В самое жаркое время многие пустынные псаммофиты сокращают испаряющую поверхность путем сбрасывания части листьев и сочных ветвей или сезонной
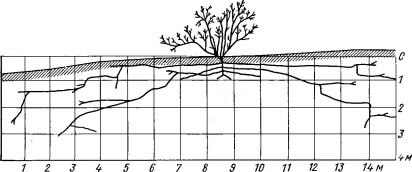
Рис. 145. Корневая система джузгуна голова Медузы — Calligonum caput medusae в барханных песках Каракумов (по Петрову М. П., 1950) 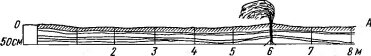
Ом 1 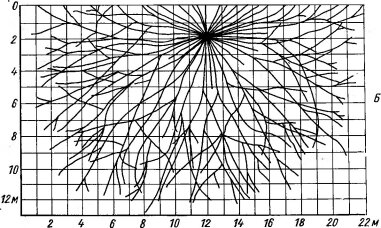
Рис. 146. Корневая система пустынного злака — Arislida karelini (по Петрову М. П., 1950). А — профиль, Б — план
смены листьев. (Ряд других приспособлений пустынных ксерофитов описан в гл. IV.)
Еще одна особенность, помогающая псаммофильным кустарникам преодолевать недостаток воды,— использование влаги глубоких горизонтов благодаря развитию мощной, глубоко проникающей корневой системы, как у крупных кустов и деревьев саксаула, почти всегда связанной с капиллярным подпором грунтовых вод.
Среди травянистых псаммофитов много эфемеров и эфемероидов, вегетирующих только ранней весной, когда в пустыне еще есть влага.
Растения-псаммофиты встречаются на песчаном субстрате и вне пустынь, в областях более умеренного климата —- по берегам морей, крупных озер, на пойменных песчаных террасах вдоль рек и т. д. Это волоснец (Elymus giganteus), цмин песчаный (Hely- chrysum arenarium), лапчатка песчаная (Potentilla arenaria) и др. Произрастая не в столь жестких условиях, как пустынные псаммофиты, эти растения не имеют выраженных специализированных черт; но и у них встречаются корневые чехлики, длинные цепкие корневища, защищающие подземные части от выдувания, а также ксероморфные черты в структуре надземных частей (например, восковой голубоватый налет на листьях волоснеца, густое серебристое опушение листьев и цветки-«бессмертники» с сухим околоцветником у цмина и т. д.)
У некоторых североамериканских псаммофитов, растущих на Тихоокеанском побережье в условиях сезонных колебаний уровня песка, отмечена интересная особенность ростовых процессов — соответствующая сезонная периодичность роста с пиком в апреле— мае, до начала подъема уровня песков.
В практике мелиорации многие псаммофиты применяют для закрепления подвижных песков. Так, способность кустарниковой ивы-шелюги (Salix acutifolia) к быстрому укоренению используется при «шелюговании» песков.
Литофиты. Это растения, обитающие на камнях, скалах, каменистых осыпях, в жизни которых преобладающую роль играют физические свойства субстрата. К этой группе принадлежат прежде всего первые (после микроорганизмов) поселенцы на скальных поверхностях и разрушающихся горных породах: сначала авто- трофные водоросли (Nostoc, Chlorella и др.), за ними накипные лишайники, плотно прирастающие к субстрату и окрашивающие скалы в разные цвета (черный, желтый, красный и т. д.), и, наконец, листоватые лишайники. Выделяя продукты метаболизма, литофильные лишайники способствуют разрушению горных пород (биологическое выветривание) и на ранних стадиях колонизации каменистых субстратов играют существенную почвообразующую роль. Процесс этот очень длительный из-за медленного роста самих лишайников (так, в горных условиях налет лишайников на скалах ежегодно увеличивается в толщину не более 0,1 мм), однако со временем на поверхности и особенно в трещинах камней накапливается слой органических остатков, на которых поселяются мхи. Под моховым покровом создается уже слой примитивной почвы, на которой поселяются и литофиты из высших растений. Используя трещины в камнях и скалах, заполненные почвой, некоторые виды способны жить на очень малом количестве субстрата (их называют хасмофитами—растениями щелей, рис. 147). Среди них виды рода камнеломка (Saxifraga), кустарники и древесные породы (можжевельник, сосна, рис. 148; в Крыму и на Кавказе — скальные формы бука, дуб скальный). Они обладают особой формой роста (ползучей, искривленной, карликовой и т. д.), связанной как с недостатком питательного субстрата, так и с жестким водным и тепловым режимом на скалах, сильными ветрами.
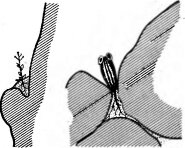
Рис. 147. Местообитания растений на скалах пустыни Намиб (по Вальтеру Г., 1968)
Так, нередко можно видеть, как побеги горных кустарников плотно прижимаются к крупным камням (рис. 149). Такие формы роста позволяют растениям использовать тепловое излучение нагретого каменного субстрата. Одна из главных особенностей растений скал и осыпей — неглубокая, но очень прочная и цепкая корневая система.
Своеобразные условия для растений-литофитов создаются на горных каменистых осыпях — новых субстратах, возникающих в результате выветривания и разрушения скал, или вторичных, образующихся под влиянием неумеренного выпаса на горных склонах и обнажения щебнистого грунта. Растения вынуждены приспосабливаться здесь не только к режимам экологических факторов,
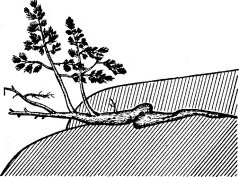
Рис. 148. Скальная форма роста сосны на гранитных скалах. Побережье Ладожского озера (по Ниценко А. А., 1951)
обусловленным каменистым субстратом, но прежде всего к подвижности осыпей. У многих видов отмечаются различные способы закрепления стеблевых частей: например, они погружены в каменистый субстрат так, что над поверхностью поднимаются лишь невысокие (несколько сантиметров) розетки листьев и соцветия. У других на небольшой глубине под поверхностью развиваются удлиненные ползучие побеги. При засыпании растения осыпей, следуя за движением субстрата, способны формировать придаточные корни и новые побеги. Корневые системы на осыпях обычно ориентированы вверх по склону: растения, подобно альпинистам, как бы «цепляются» за опору вверху. Часто развиваются контрактильные (втягивающие) корни.
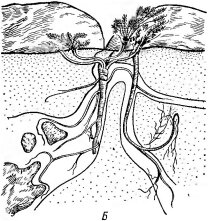
Пронизывая осыпь сеткой корневищ и корней, расте- ния-литофиты могут замедлить и приостановить ее движение, давая возможность селиться видам с надземными побегами и глубокими корневыми системами, которые хорошо удерживают субстрат («плотинные» растения). На следующем этапе происходит накопление мелкозема и растительных остатков — начинается почвообразовательный процесс. Впоследствии на таких остановленных осыпях могут появиться и группы дерновин- ных растений — очаги будущих высокогорных лугов.
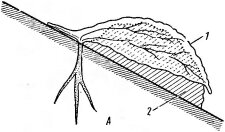
В роли случайных лито- фитов оказываются растения самых разных видов, поселяющиеся на старых каменных строениях, старинных крепостях, каменных оградах и т. д. В расщелинах
Рис. 149. Остролодочник углубленный— между камнями, где скапли- Oxytropis immersa — пример высокогорного ВЭЮТСЯ ПЫЛЬ И МЄЛКОЗЄМ, НЄ- растения-литофита. Западный Тянь-Шань редко МОЖНО видеть травя- (по Культиасову И. М.. 1955). А — побеги НИСТЫе многолетники и даже „О..рх„о™ сисе- деревца.
/ — побеги, 2 — камень В ПОСЛЄДНИЄ ГОДЫ ЭКОЛО
ГИЯ многих красивоцвету- щих литофитов привлекла к себе внимание в связи с распространением «альпийского» стиля в цветоводстве (устройство альпинариев) и введением дикорастущих литофитов в культуру.
Еще по теме ВЛИЯНИЕ ЗАСОЛЕНИЯ НА РАСТЕНИЯ:
- Кислотные дожди и их влияние на растения
- О влиянии свойств почв на стойкость растений.
- ВЛИЯНИЕ ХОЛОДА НА РАСТЕНИЯ И ПРИСПОСОБЛЕНИЯ К НЕМУ
- ОСОБЕННОСТИ ВЛИЯНИЯ БИОТИЧЕСКИХ ФАКТОРОВ. ЭКОЛОГИЧЕСКИЕ СВЯЗИ НАСЕКОМЫХ С РАСТЕНИЯМИ
- ВЛИЯНИЕ ЖИВОТНЫХ НА ХИМИЧЕСКИЕ СВОЙСТВА ПОЧВЫ И ПРОДУКТИВНОСТЬ РАСТЕНИЙ
- МНОГОЛЕТНИЕ ЦИКЛИЧЕСКИЕ ИЗМЕНЕНИЯ В СРЕДЕ И ИХ ВЛИЯНИЕ НА ЖИЗНЬ РАСТЕНИЙ
- ВЛИЯНИЕ КАЛИЙНО-ФОСФАТНОГО ФОНА НА ИСПОЛЬЗОВАНИЕ РАСТЕНИЯМИ АММИАЧНЫХ И НИТРАТНЫХ ФОРМ АЗОТА[18]
- ВЛИЯНИЕ ВНЕШНЕЙ СРЕДЫ НА УСЛОВИЯ ПИТАНИЯ РАСТЕНИЙ И ЭФФЕКТИВНОСТЬ УДОБРЕНИЙ
- Влияние доз и времени внесения калия в условиях аммиачного и нитратного питания растений на их урожай и качество продукции
- РАСТЕНИЯ-ЧАСЫ, РАСТЕНИЯ-СИНОПТИКИ
- КОСВЕННЫЕ ВЛИЯНИЯ
- Влияние влажности на насекомых
- Влияние гиподинамии
- Положительное влияние
- прямые влияния
- ВЛИЯНИЕ АНТРОПОГЕННЫХ ФАКТОРОВ НА НАСЕКОМЫХ
- ВЛИЯНИЕ ОНДАТРЫ НА BOX НУЮ РАСТИТЕЛБНОСТБ
- 3.3 ХИМИЧЕСКИЙ СОСТАВ ПОЧВЫ И ЕГО ВЛИЯНИЕ НА ЗДОРОВЬЕ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ