ОСОБЕННОСТИ ВЛИЯНИЯ БИОТИЧЕСКИХ ФАКТОРОВ. ЭКОЛОГИЧЕСКИЕ СВЯЗИ НАСЕКОМЫХ С РАСТЕНИЯМИ
Зависимость влияния биотических факторов от плотности популяций. Одностороннее приспособление насекомых к физической среде и двусторонние адаптации сочленов биоценозов. Связи насекомых с растениями.
Вред, наносимый растениям насекомыми. Классификация повреждений растений. Перенос насекомыми возбудителей‘ заболеваний растений и поражаемость насекомых грибными и бактериальными организмами. Насекомоядные растения. Защитные приспособления растений к повреждениям насекомыми и защитные реакции насекомых по отношению к патогенным растительным организмам. Приспособление насекомых к защитным реакциям растений. Опылительная роль насекомых и взаимные адаптации насекомых и растений, обеспечивающие энтомофилию. Взаимообусловленность эволюции насекомых и Цветковых растений. Симбиоз. Роль насекомых в расселении растений и растений в расселении насекомых. Роль растений в качестве убежищ для насекомых. Взаимная зависимость численности насекомых и растений в биоценозе и распределение их популяций на территории биотопа.Биотические факторы среды в отличие от абиотических по своему действию на насекомых имеют некоторые специфические особенности.
Факторы абиотические — температура, влажность, осадки и т. д. — действуют, как правило, на всю популяцию насекомых более или менее однородно, возможны лишь сравнительно небольшие индивидуальные и групповые отклонения.
Влияние же биоценотических связей в большинстве случаев затрагивает только больший или меньший процент особей популяции. Лишь в очень редких случаях
окажутся заражены каким-либо паразитом или съедены хищником все особи данной популяции. Почти никогда не бывает гибели всей популяции от какого-либо инфекционного заболевания. Только немногие .биотические факторы носят иной характер. Например, некоторые симбиотические отношения, в частности отношения с микроорганизмами, населяющими кишечник и помогающими усваивать те или другие вещества пищи (растительную клетчатку и др.), впрочем, эти симбиотические отношения изучены недостаточно.
Степень влияния биотических факторов на популяцию зависит обычно от численности особей на единицу площади, т. е. от плотности популяции. Если температура действует на все особи популяции независимо от того, находится ли на 100 м2 площади 10 или 10 000 экземпляров данного вида, то для паразитов это далеко не безразлично. Чем гуще будет население хозяина на данной территории, тем легче паразитам отыскать хозяина. При большем скоплении особей на единице площади, с большей быстротой распространяются от одного насекомого к другому возбудители инфекционных заболеваний. Конкуренция между насекомыми, нуждающимися в одинаковых кормовых растениях, имеет тем меньшее значение, чем больше в данном биотоне таких растений, и тем большее значение, чем плотнее, гуще будет население' этих насекомых в биотопе.
В связях насекомых с абиотической средой имеется одностороннее приспособление насекомых к среде. Это, конечно, не значит, что насекомые не влияют на окружающую абиотическую среду. Из предыдущих глав об обмене веществ и энергетическом обмене насекомых со средой это видно достаточно ясно. Однако влияние насекомых на физическую среду их обитания (уж не говоря О' том, что это влияние не распространяется сколько-нибудь заметно далее микроклиматических изменений на небольших участках) само по себе является в большинстве случаев приспособлением к физической среде.
В биотических же связях, связях между организмами, типично взаимное приспособление, двухсторонняя адаптация организмов друг к другу. Если хищник или паразит стремятся отыскать свою жертву, то жертва стремится спастись от паразита или хищника. Если насекомое приспособляется к цветам растений для питания их нектаром, то и растение, для которого посещение насекомых полезно, приспособляется к опылению насекомым, и даже самое выделение нектара произошло как адаптивное к посещению насекомых явление. Если возбудители заболеваний вырабатывают (в историческом аспекте) способы, наиболее обеспечивающие наилучшие для них возможности поражения насекомых, то и у поражаемых насекомых в качестве ответа создаются пути и способы, ведущие к тому, чтобы избежать заболевания.
Двусторонняя адаптация организмов проявляется очень сложно, тем более, что, кроме прямых связей между организмами, существуют и косвенные; взаимоотношения не ограничиваются только двумя видами, но захватывают и весь биоценоз. Кроме того, весьма важно подчеркнуть, что проявление выработанных насекомыми адаптивных черт в большой мере зависит и от разного сочетания факторов абиотической среды. Влияние на численность популяций биотических и абиотических факторов среды носит комплексный характер, как это будет показано в следующей главе.
Все многообразие изменяющихся условий среды ведет к значительным колебаниям численности всех видов насекомых в любом биотопе.
Непонимание диалектических связей между средой и организмом повело к появлению идеалистических теорий о якобы существующем в природе биоценоти- ческом равновесии, о более или менее 'полном саморегулировании численности отдельных видов на примерно одинаковом уровне. Было высказано предложение рассматривать биоценоз как организм высшего порядка, как физиологическую единицу жизни и т. д. (Тинеман, 1918; Клементс, 1928, 1939; Жерард, 1942; Эмерсон, 1939, 1949, 1950 и др.). Не менее порочны противоположные попытки оторвать организмы от среды, обосновывать «равновесие» в размножении отдельных видов «сопротивлением среды» (Чепман, ] 925, 1928; Бремер, 1928 и др.).
Организм и окружающая его среда представляют собой единство, но единство1 это не равнозначно отдельному организму, хотя бы уже по одному тому, что между организмами существуют очень своеобразные внутривидовые и межвидовые связи.
Выше говорилось, что в основе биотических отношений между видами в биоценозе лежат пищевые цепи, которые связываются в циклы питания, и что каждая из этих цепей, как правило, начинается с растения. В пище насекомых, как и всех животных, должны быть белки, жиры и углеводы, поэтому они питаются лишь органическими веществами. Источником этих веществ или непосредственно, или через* других животных — фитофагов, могут быть только растения.
Понятно, таким образом, что растения должны играть большую прямую или косвенную роль в жизни любого насекомого.Экологические связи насекомых с растениями выражаются, в основном, в питании за счет различных частей растений, косвенно также в питании за счет животных — фитофагов и в паразитизме, а в редких случаях в хищничестве, некоторых растительных организмов на насекомых. Во многих случаях растения используются насекомыми в качестве убежища от неблагоприятных метеорологических условий или укрытия от их естественных врагов (паразитов и хищников). Некоторые насекомые могут расселяться по территории с семенами и опавшими частями растений. Известны случаи симбиоза насекомых с одноклеточными грибами и бактериями, живущими в полости тела насекомых.
Значение насекомых для растений определяется питанием на них' насекомых, влиянием насекомых на численность и жизненность других растительноядных животных, переносом насекомыми возбудителей заболеваний растений, влиянием насекомых на почвенные процессы (аэрацию, гумификацию и проч., о чем будет сказано в главе 12). Некоторые растительные организмы, в свою очередь, развиваются непосредственно на насекомых, таковы, в частности, энтомофторовые и некоторые одноклеточные грибы. Многие бактерии также развиваются за счет насекомых. Есть и высшие насекомоядные растения, способные усваивать белковые вещества. Насекомые помогают расселению растений по территории. Растения и насекомые взаимно обусловливают их географическое распространение.
Питание насекомых на растениях далеко не во всех случаях относится к антагонистическим отношениям, особенно велико взаимное положительное значение питания насекомых — опылителей, в данном случае создаются симбиотические связи.
Насколько может быть велик вред, наносимый растениям питающимися на них насекомыми, видно из огромного количества наблюдений, учетов и специальных исследований, опубликованных специалистами по сельскохозяйственной и лесной энтомологии. Приведем некоторые примеры.
Пять экземпляров клопа вредной черепашки (Eurygdster integriceps Put.) во взрослом состоянии могут уничтожить весь урожай пшеницы или ячменя в период выхода в трубку на площади 0,25 м2 (Родд, Гуссаковский и Антонова, 1933). Всхожесть зерна, полученного с зараженных черепашкой полей, снижается до 50%. Свекловичная (бобовая) тля (Aphis fabae Scop.) может снизить сахаристость корней свеклы до 69%, а их вес —до 7,4%. На семенниках свеклы эта тля иногда губит урожай семян полностью.Пораженные гороховой зерновкой (Bruchus piso- rum L.) семена гороха теряют до 75% всхожести. Просяная мушка (Stenodiplodis panici Rohd.) при массовом размножении может погубить весь урожай проса. Повреждения прибрежной мушки (Ephydra macellaria Egg.) в Средней Азии иногда вызывают необходимость пересевов риса. Взрослый свекловичный долгоносик (Bothynoderes punctiventris Germ.) на Украине иногда полностью губит всходы свеклы на больших площадях. Гусеницы беленной совки (Chloridect pettigera Schiff.) уничтожают в Средней Азии до 50—85% бутонов и цветков сафлора. Отмечены случаи гибели до 97% растений на посевах кунжута от кунжутной златки (Acmaeodera batlioni Gangl).
Полная гибель молодых деревьев персиков наблюдается иногда от оранжерейной тли (Myzodes persicae Sulz.), молодых деревьев яблонь — от кровяной яблоневой тли (Eriosoma lanigerum Hausm,). Плодовые декоративные и лесные деревья частично или полностью оголяются от листьев листогрызущими видами вредных насекомых, например, яблоневой молью (Hyponomeuta malinellus Zel.), непарным шелкопрядом (Porthetria dispar L.), тутовой пяденицей (Apocheima cinerarias
Ersch.) и многими другими, а это ведет к резкому снижению урожая плодов и семян, Так, по данным Б. Г. Шуровенкова (1953), яблоня сорта Апорт в Ал- маатинской области при объедании листьев яблоневой молью на 25—100% теряет урожай плодор в этом году на 16,4—71,1%, а ослабленные деревья на следующий год после повреждения дают урожай по сравнению с неповрежденными деревьями на 21 —100% ниже.
Средний коэффициент вредности кукурузного мотылька (Pyrausta nubilalis Hb.) на кукурузе равен 15,25, средний коэффициент вредности листового люцернового слоника (Phytonomus variabilis Hbst.) в Средней Азии равен 65 [10] .(Яхонтов, 1953). Общеизвестно, что во многих странах огромный вред йаносят восточная азиатская саранча {Locusta migratorіа типі- lensis Меуеп.), перелетная саранча (L. migratoria migrator іа L.), розовый коробочный червь (Pectinophora gos- sypiella Saund.), рисовые огневки (Schoenobius incertel- lus Walk, и Chilo simplex Butler.), пшеничный комарик (Sitodiplosis mosellana Gehin.), луговая совка (Cirphis unipuncta Haworth.), хлопковая тля (Aphis gossypit Glov.) и многие другие насекомые.
Общие потери урожая культурных растений из-за питания вредных насекомых достигают огромных размеров. Так, например, в США в 1946 г. было уничтожено 15,1% урожая хлопчатника, в 1950 т. эти потери исчислялись в 908 млн. долларов. Ежегодные потери от всех вредителей в сельском хозяйстве и лесоводстве достигают в США в последнее .время в среднем около 4 млрд. долларов.
Правда, все это относится преимущественно ко вторичным биотопам, где создаются для вредных насекомых, нередко, особо благоприятствующие размножению условия среды, особенно в смысле обилия пригодных питающих растений. Однако и в свободной природе иногда наблюдается огромный вред, наносимый растениям насекомыми. Так, например, заросли дикой яблони в горах Южного Казахстана, Южной Киргизии и Узбекистана иногда почти нацело оголяются яблоневой молью {Hyponomeuta malinetlus Zell.); автор наблюдал ,в горах Южного Казахстана дикие заросли кустарника Cotoneaster Medik., совершенно объеденные жуками Litta menetriesi Fid.; азиатская саранча {Locusta migratoria L.) часто на больших площадях по- берегам рек и озер, в местах ее резерваций, объедает листья тростника Phragmites communis Trin. и т. д.
Вред, наносимый растениям питающимися на них насекомыми, состоит или в обгрызании их органов (почек, листьев, бутонов, цветков, плодов, стволов, стеблей, ветвей, корневой шейки, корней и т. д.), или в высасывании растительных соков из этих органов. В результате повреждений нарушается нормальное поступление питательных веществ и воды из почвы, нарушаются дыхательные процессы, снижаются темпы фотосинтеза из-за потери хлорофилоносных тканей. Введенные при сосании в ткани растений выделения слюнных желез насекомых нередко разрушают крахмальные зерна и вызывают ненормальное накопление в клетках глюкозы и т. д. В результате питания насекомых происходит задержка в развитии растений. Например, согласно проведенной автором работы, гибель верхушечной почки всхода хлопчатника из-за сосания табачного трипса (Thrips tabaci Lind.) или хлопковый тли (Aphis gossypii Glov.) задерживает развитие всхода хлопчатника в Узбекистане в среднем на четыре дня, а гибель от сосания этих вредителей вновь образующейся почки сбоку семенодольных листьев приводит в среднем к двухнедельному отставанию, растения в развитии (Яхонтов, 1929).
Насекомые с грызущим ротовым аппаратом при питании отгрызают отдельные участки растений; при высасывании насекомыми соков отдельные поврежденные участки растительных тканей отмирают и дефор миру- ются. Ослабленные питанием насекомых растения резко снижают урожай плодов и семян, они менее устойчивы к заболеваниям, часто (подвергаются нападению вторичных вредителей. Вторичными вредителями называют таких вредителей, которые на здоровые, крепкие растения, как правило, не нападают, для них необходимо предварительное ослабление растений; например, многие короеды (Ipidae), златки (Buprestidae), некоторые жуки усачи (Cerambycidae).
Характер повреждения растений насекомыми при питании классифицируют следующим образом.
Различают повреждения частей растений (бутонов, завязей, плодов, корней, стеблей и стволов, ветвей, листовых и цветочных почек) наружное и внутреннее. Примером внутреннего повреждения бутонов, завязей и плодов моЖет служить питание розового коробочного червя (Peciinophora gossypiella Saund.) или гусениц хлопковой совки (Chloridea obsoleta F.) (рис. 58) на хлопчатнике. Группа бабочек, гусеницы которых повреждают плоды фруктовых и ореховых деревьев, носит даже специальное название «плодожорок», таковы, например, яблочные плодожорки (Carpocapsa pomonella L., Carposina sasakii Mats.), восточная плодожорка Lctspeyresia molesta Busck.), сливяная плодожорка (L. funebrana Tr.), ореховая плодожорка (Sarrothrypus musculana Ersch.). Внутреннее повреждение почек на плодовых деревьях производят, например, гусеницы чех- лико вой моли (Coleophora hemerobiola Fil.) и вишневый слоник (Rhynchites auratus Scop.).
Наружное повреждение бутонов, почек, завязей, плодов производят, например, щитовки (Coccodea), тли (Aphidodea), некоторые клопы (Hemiptera), многие гусеницы бабочек. На верхних частях стеблей кенафа из- за сосания клопов иногда вообще не остается ни одного бутона или коробочки. -
Внутреннее повреждение корней растений производят личинки некоторых златок (Buprestidae), личинки клубеньковых слоников (Sitona sp. sp.) на бобовых растениях и др.
Наружное повреждение корней свойственно некоторым тлям (Trifidaphis phaseoli Pass.; Brachynnguis anu- raphoides Nev., Forda triviatis Pass, и др.), .личинкам многих жуков (например, хрущей — Melolonthinae, чер-
Рис. 58. Коробочка хлопчатника, поврежденная гусеницей хлопковой совки (ориг.; зарисовка Короби- цина)
нотелок — Tenebrionidae, щелкунов — Elateridae и т. д.).
Внутреннее повреждение стеблей и стволов растений производят, например, личинки многих жуков златок (Buprestidae), усачей (Cerambycidae) (рис. 59), короедов (Ipidae), личинки хлебного пилильщика (Cephus pygmaeus L.), гусеницы бабочек стеклянниц (Sesiidae) и древоточцев (Cossidae), стеблевого (кукурузного) мотылька (Pyrausta nubilalis Hb.), рисовых огневок (Chilo simplex Butler, Schoenobius incertetlus Walk.).
Наружное .повреждение стволов, стеблей и ветвей растений характерно для имаго сафлорного слоника (Mesogroicus petraeus Faust.), большой (стволовой) пер-
Рис. 59. Поперечный срез ствола, погибшего от повреждения усачом Aeolestes sar- ta Sols, (ориг.)
сиковой тли (Pterochloroides persicae Choi.) и некоторых других тлей, очень многих кокцид (Coccodea) и т. д.
Если стебель, ветку или корень насекомые перегрызают поперек или вокруг в виде кольца, то такое повреждение называют подгрызанием, или п е р е- гры заниєм. Таковы, в частности, повреждения гусениц совок подсемейств Agrotinae, личинок жука Heto- phorus micans Fald., взрослых маек (Meloe sp. sp.), некоторых видов взрослых короедов (Ipidae), как, например, Scolytus mali Bechst. и S. rugulosus Ratz.
Повреждение листьев бывает сплошное, когда насекомое поедает весь лист; краевое — когда насекомое выедает-с краев листа участки того или иного размера; дырчатое — когда насекомое на листе выедает сквозные дыры; окошчатое — когда насекомое выгрызает на плоскости листа более или менее мелкие участки, оставляя не тронутой верхнюю или нижнюю кожицу; скел етирование — если насекомое съе-
дает мякоть и кожицу листа, оставляя нетронутыми жилки; минирование — когда насекомое, проникши через эпидермис в одном месте, выедает паренхиму листа. По форме различают мины: звездчатые,
змеевидные, линейные, пятно образ аы е, т р у б к о о б р а з н ы е, а в зависимости от поврежден- ности всей мякоти листа или только его верхней палисадной ткани, или нижней губчатой ткани, мины называют двухсторонними, верхнє сторонними или нижнє сторонни ми.
Когда в результате сосания насекомых на листе образуются небольшие пятнышки желтого, белого, бурого или черного цвета, повреждение называется пятнистым. Если листья для питания скрепляются насекомыми вместе паутинкой, повреждения называются паутинными гнездам и, при свертывании их в трубку — трубчатым и; если происходит сморщивание или скручивание,— повреждения называют деформацией листьев.
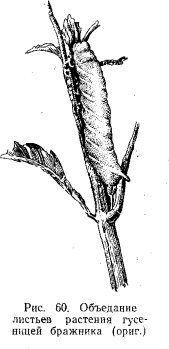
Примерами сплошного повреждения листьев может служить съедание их целиком гусеницами бабочек бражников (Sphingidae) в старших возрастах (рис. 60) или другими крупными гусеницами, а также некоторыми саранчовыми (Acridodea) во взрослой фазе или их личинками в старших возрастах. ¦
В качестве примеров насекомых, производящих краевые повреждения листьев, можно назвать листовых древесных слоников (Polydrosus Germ.), гусениц бабочек (Lepidoptera) самых различных семейств, саранчовых (Acridodea), сверчков (Gryllodea), кузнечиков (Tettigoniidae) и др. Но характер питания, а следовательно, и тип повреждения листьев в разных возрастах личинок прямокрылых насекомых (Orthoptera) и гусениц бабочек может изменяться. Те же виды, которые в последних возрастах производят краевое или сплошное повреждения листьев, в молодом возрасте нередко минируют, скелетируют листья или производят дырчатые повреждения. Таковы, например, прусик (Calliptamus italicus L.), или гусеницы капустной белянки (Pieris brassІсае L.) и многие др.
Очень характерны и своеобразны краевые повреждения листьев пчелами из рода Megachile Latr., выгрызающими правильные овалы, полуовалы или полукруги (рис. 61), но в этом случае отгрызенные части листа используются не для питания, а для построения ячеек гнезд, устраиваемых пчелами в почвенных норках или в пустотах стеблей растений.
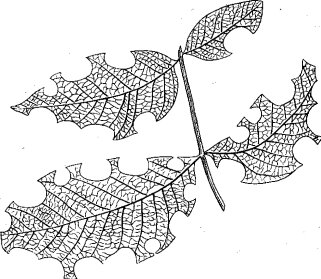
Рис. 61. Лист грецкого ореха, поврежденный пчелами рода Megachile Latr. (ориг.)
Дырчатые повреждения листьев производят, напри мер, гусеницы средних и старших возрастов совки ка? радрины (Laphygma exigua Hb.), гусеницы урюковой совки (Catymnia sub til is Stgr.) и капустной моли (Plu- telta maculipennis Curt.) в средних и старших возрастах.
Окошчатые повреждения листьев свойственны питанию взрослых мелких листоедов из родов Phytlotreta, Foudr. и Chaetonema Steph., а также многих других насекомых. Иногда окошчатое повреждение впоследствии превращается в мелкодырчатое, в результате разрушения и выпадения оставленной при питании тонкой эпидермальной пленки листа (рис. 62).
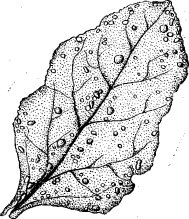
Рис. 62. Лист свеклы, поврежденный листоедом Chaetonema bre- viuscala FaU. (ориг.)
Скелетирование характерно для многих гусениц бабочек в младших возрастах, например, для непарного шелкопряда (Porthetria dispar L.), карадрины (Laphygma exigua Hb.), для многих взрослых жуков листоедов и их личинок, например, для тополевого листоеда (Melasoma рори- ІІ L.) (рис. 63), рапсового листоеда (Entomoscelis adonidis Pall.), пьявицы (Lema melanopus L.), ложногу- сениц пилильщика Caliroa cerasi L/и др.
В качестве примеров минирующих насекомых можно
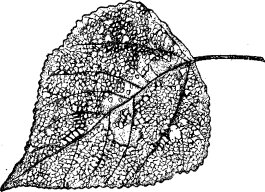
Рис. 63. Лист тополя, скелетированный тополевым листоедом (ориг.)
указать на гусениц первого возраста яблоневой моли (Hyponomeuta malinellus Zell.), гусениц первого возраста листовой вертуньи (Recurvaria nanella Schiff.), гусениц чехликовой моли (Coleophora hemerobiola Fill.), кружковой моли (Cemiostoma scitella Zell.), бобовой выемчатой моли {Anacampsis cmthylidetla Hb.), личинок мух родов Phytomyza Fall., Hydrellia R. D., личинок ґ свекловичной мухи (Pegomyia hyosciami Panz.). Этот7 тип повреждений свойствен преимущественно представителям отрядов Lepidoptera и Diptera, реже Coleoptera.
Пятнистость листьев производится сосущими насекомыми. Так, например, в результате сосания табачного трипеа (Thrips tabaci Lind.) образуются около жилок мелкие серебристые пятнышки. От сосания клопа вредной черепашки (Eurygaster integriceps Put.) остаются белые, потом буреющие пятнышки, бурые круглые пятнышки остаются после питания люцернового клопа (.Adetphocoris tineolatus Goeze.), светлые многочисленные пятнышки сверху листьев яблони и груши оставляют клопы Stephanitis oshanini Vas., S. pyrі F. (с нижней стороны поврежденных этими клопами листьев появляются многочисленные мелкие черные ПЯТНЫШКИ — не результат сосания, а экскременты).
Примерами паутинных гнезд могут служить скрепленные паутинками пучки листьев, внутри которых питаются поодиночке средне- и старшевозрастные гусеницы почковой вертуньи (Tmetocera oeellana F.); листовой вертуньи (Recurvaria nanella Schiff.); крупные сплетения паутины вокруг пучков листьев или вокруг целых ветвей, в которых коллективно держатся гусеницы яблоневой и плодовой молей (Hyponomeuta malinellus Zell, и Н. padellus L.), а позднее находятся их коконы; устраиваемые на зиму гусеницами боярышницы (Aporia crataegi L.) укрытия из листьев, скрепленных паутинкой и прикрепленных паутиной к ветвям деревьев; паутинные сплетения, устраиваемые на листьях маревых (Chenopodiaceae) и некоторых других растений гусеницами туркестанского свекловичного мотылька (Loxostege nudalis L.). Паутинные гнезда на растениях устраиваются в основном гусеницами бабочек (Lepidoptera); в тропических и субтропических районах скрепляют листья паутинкой на деревьях некоторые муравьи (Fprmicidae), как, например,
Oecophila smaragdina Fabr., которые устраивают среди них свои колонии, но при этом они не питаются растениями.
Свертывают листья в виде трубок жуки трубковерты, например, Rhynchites betulae L. на березе (личинки трубковертов питаются свернутыми листьями внутри трубок, сделанных взрослыми жуками); гусеницы многих видов бабочек семейства листоверток (Tortricidae), получивших такое же характерное название, как и жуки трубковерты. Трубкообразные свертывания листьев происходят в результате питания некоторых тлей (например, спирально окрученные трубки образует на. яблонях тля Yezabura affinis Mordv. : или из верхушечного листа ячменя тля Brachycolus noxius Mordv.).
Деформации в виде неправильного скручивания и сморщивания листьев производят многие сосущие на растениях насекомые: табачный трипе (Thrips tab асі Lind.) на листьях хлопчатника, тли (Aphidodea) на различных растениях, иногда цикады (Auchenorhyncha), как, например, пен ниц a Aphrophora spumaria L.
Деформация и пятнистые повреждения молодых листьев в дальнейшем ведут к разрывам листовой пластинки вследствие отмирания отдельных участков листа,, который продолжает увеличиваться в размерах. Это наблюдается, например, при питании табачного трипса (Thrips tabaci Lind.), некоторых тлей (Aphidodea), цикад (Auchenorhyncha). Края разрывов впоследствии зарубцовываются; иногда разрывы имеют сложную зубчатую форму, если отмирание тканей происходило во многих точках. Такие повреждения также относятся по типу к деформациям.
Кроме названных типов повреждений, на листьях, а также на ветвях, стеблях и корнях, в результате раздражающего действия слюны, проникающей в. ткани растений при сосании насекомых и соответствующей реакции растения, иногда возникают опухоли различной величины и формы, характерные Для вида повреждающего насекомого; такие разрастания тканей растения называют галлами.
Галлы бывают сплошными или полыми, в последнем случае насекомые находятся внутри галлов и покидают их только' после образования на них трещин. Сплошные галлы, "имеющие крупную величину,
иногда называют раковыми опухолями, хотя никакой аналогии с раковыми разрастаниями тканей у людей и животных здесь не имеется. Примером сплошных галлов могут служить опухоли, вызываемые на стволах, ветвях и на верхней части корней яблонь сосанием кровяной яблоневой тли (Eriosoma lanigerum Наизт.). В качестве примеров полых галлов можно назвать мешковидные образования на жилках и черешках листьев фисташки, возникающие в результате сосания тли Slavum lentiscoides Mordv. (рис. 64), опухоли, вызываемые на листьях тополей тлями рода Pemphigus Hartig., огромные (иногда крупнее кулака взрослого человека) вздутия на листьях вязо'в, образуе-
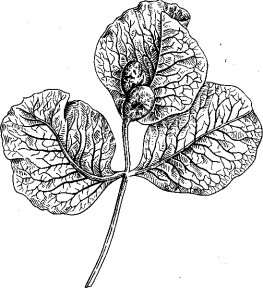
Рис. 64. Галлы тли Slavum lentiscoides Mordv. на листе фисташки (ориг.)
мые тлями Eriosoma lanuginosum Hartig., и вздутия меньшей величины, образуемые на вязах тлями Е. patchiae Вот. Вздутия на соломинах злаков, состоящие из нескольких камер, внутри которых находится по одной личинке галловой толстоножки (HarmoUta rossica Rim. Kors.), или опухоли, вызываемые на ветвях деревьев и кустарников гусеницами стеклянниц (Aege- riidae).
Между полыми и сплошными галлами существуют переходы. Например, галлы, в которых вредитель находится короткое время, а затем обитает снаружи галла,таковы галлы пилильщиков рода Pontania Costa на иве. Некоторые галлы, вызываемые орехотворками (сем. Cynlpi- dae) или мушками Itonididae, хотя личинки находятся и внутри них, имеют очень небольшие полости при очень толстых стенках галла, так что полыми они могут быть названы лишь очень условно.
Насекомые вредят растениям не только своим питанием, известны и иного характера повреждения. В частности, вред может происходить в результате поранения тканей растений яйцекладом самок при яйцекладке. Крупные хлопковые цикады {Cicadatra ochreata Mel. и С. querula Pall.) по границам оазисов Средней Азии производят в отдельные годы массовое усыхание вершин растений, перерезая при яйцекладке проводящие сосудистые пучки стеблей (Яхонтов, 1929).
Подобные же повреждения производят сверчки Oecanthus pellucens Scop, и О. turardcus Uv., откладывая яйца в стебли и ветви различных культурных и дикорастущих растений, следствием чего является засыхание вершин или опадение на них бутонов, цветков или плодов.
Очевидно, должна оказывать отрицательное влияние и яйцекладка внутрь растительных тканей других насекомых — трипсов (Thysanoptera), мелких видов цикад (Auchenorhyncha), жуков (Coleoptara) и проч. При яйцекладке, в результате раздражения растительных тканей выделениями придаточных желез половой системы самок, иногда образуются галлы, подобные галлам, вызванным питанием сосущих насекомых, например, небольшие вздутия образуются над яйцами некоторых трипсов — одуванчикового (Thrips taraxaci Yakh;), трип- са фрица (Taeniothrips frici Uzel.), откладывающих яйца в эпителий обертки цветущих корзинок одуванчиков (Яхонтов, 1953), и др. Галлы некоторых орехотворок (Cynipidae) и пилильщиков (Tenthredinidae) также возникают не только в результате питания их личинок, но и в результате выделений придаточных желез половой системы, вводимых в растительные ткани при яйцекладке.
Выше упоминалось о том, что на листьях яблонь и груш остаются с нижней стороны очень многочисленные пятнышки экскрементов клопов (Stephanitis pyri F. и S. oshanini Vas.), эти экскременты закрывают большое количество устьиц на листьях, что отрицательно действует на газообмен деревьев. Подобный вред могут наносить и некоторые другие обитающие на растениях насекомые.
Перенос возбудителей заболеваний растений насекомыми совершается чаще всего во время их питания. В последние десятилетия появилась обширная литература по вопросу о передаче растениям вирусных заболеваний. В отношении многих вирусных болезней передача вируса другим путем, кроме сосущих насекомых, впрыскивающих вирус со слюной во время питания, невозможна. Вирусная болезнь табака «рябуха» передается только тлями (Aphidodea) и табачным трипсом (Thrips tabaci Lind.), курчавость листьев картофеля — тлями Myzodes persicae Sulz. и Macrosiphum solantfoW Ashm.; болезнь «мозаика свеклы» распространяется свекловичным клопом (Poeciloscytus cognatus Fieb.). Переносчиком вируса, вызывающего болезнь курчавость листьев хлопчатника, является хлопковая тля (Aphis gossypii Glov.), вируса, вызывающего курчавость листьев сахарной свеклы, — цикада Eutettix tenella Baker. Вирус мелкоплодности вишни переносят цикады Scaphytopius acutus Say. и некоторые др.
Насекомые могут способствовать также заболеванию растений грибными и бактериальными болезнями. Например, заражение плодов различных тропических растений грибом Nematospora coryli Pegl. происходит при помощи, полужестокрылых насекомых (Hemiptera), особенно клопа Nezara viridula L.; заболевание хлопковых коробочек розовой гнилью вызывается хлопковой совкой (Chloridea obsoteta F.), повреждающей коробочки. Массовая гибель карагачей (вязов) в некоторых местах происходит в результате голландской болезни, которая передается от дерева к дереву при помощи жуков заболонников (Scolitinae). Плодовая гниль яблок и груш, вызываемая грибом Monilia Person, может распространяться долгоносиками роДа Rh.ynch.ites Schneid.; люцерновый клоп (Adelphocoris tineolatus Goeze.) способен переносить бактериозы хлопковых коробочек и т. д.
Некоторые бактерии (например, Bacterium tracheip- hilus Smith., вызывающая увядание огурцов) могут заражать растения только тогда, когда они пройдут через организм насекомого и будут введены в ткани растений со слюной этого насекомого.
Согласно учению академика Е, Н. Павловского (1939, 1946 и др.), относящемуся, правда, к переносчикам инфекционных болезней людей и домашних животных, но имеющему широкое биологическое значение, существуют природные очаги трансмиссивных заболеваний. Такими очагами являются места с исторически сложившимся комплексом возбудителей заболеваний, их переносчиков и заражаемых хозяев. Все эти компоненты биоценозов взаимно приспособлены друг к другу, они имеют синхронное развитие, хозяева и переносчики имеют соответствующую подвижность и численность, обеспечивающие постоянные контакты. В случае, если переносчиками заболевания является несколько видов животных, возможности постоянства очага увеличиваются; но, как правило, имеется основной, наиболее массовый вид переносчика заболеваний. Численность как хозяев, гак и переносчиков их заболеваний и самих возбудителей заболеваний очень зависит от условий среды.
Примерами заболеваний насекомых грибными болезнями могут служить упомянутые в главе 7 эпизоотии, вызываемые эн то мофторовьгм грибом Tarichtum phyto- nomi Jacz., среди личинок листового люцернового слоника (Phytonomus variabitis Hbst.) и Empusa aulicae Reich. — среди гусениц златогузки (Nygmia phaeo- r'rhoea L.). Очень известны осенние эпизоотии среди комнатных мух (Musca domestic a L.), вызываемые грибом Empusa muscae Cohn.; массовую гибель саранчовых (Acridodea) вызывает иногда гриб Е. grylli Nowak. В дождливые сезоны в Индии щитовки Lecanium viridis Green, в массе гибнут от гриба Cephalosporium ІесапІІ Zimm. Яблоневая медяница (Psylla malt Schm), поражается грибом Entomophthora sphaerosperrna Fres. (рис. 65), свекловичный долгоносик (Bothynoderes
punctiventris Germ.)—зеленой мюскар- диной (Metarrhizium anisopliae Sor.) (рис. 66.).

Рис. 65. Яблоневая медяница, погубленная грибом Еп- tomophthora spha- erosperma Fres (по Еремеевой)
He меньшее значение имеют бактериальные заболевания насекомых, например, бациллы Д’Ерелля (Соссо- bacillus acridiorum D’Her.), которые пытались одно время использовать в борьбе с саранчовыми насекомыми. Бактерия Bacterium galleriae Met. поражает многих гусениц бабочек; от бактерий чудесной палочки (В. prodigio- sum Ehr.) в массе погибают многие насекомые. О грибных и бактериальных заболеваниях насекомых имеются литературные сводки Штейнхауза (1950, 1952), Евлаховой и Швецовой (1953).
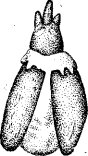
Рис. 66. Свекловичный долгоносик, погибший от заражения зеленой мюскардиной (по Евлаховой и . Швецовой)
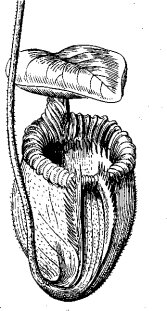
В качестве примера насекомоядных растений можно назвать болотное растение росянку (Drosea), улавливающее насекомых клейкими выделениями многочисленных желез листа, эти выделения содержат пищеварительные ферменты. Тропическое насекомоядное растение Nepenthes villosa улавливает насекомых при помощи кувшинообразного листа (рис. 67). «Кувшин», при попадании в него насекомого, закры- вается другой частью листа, играющей роль крышки. Во внутренних стенках. листа имеются железы, выделяющие пищеварительные соки.
Как у насекомых по отношению к болезнетворным растительным организмам, так и у растений по отношению к нападению вредящих им насекомых, а также передаваемых ими заболеваний, должны вырабатываться защитные, приспособительные реакции, хотя этот вопрос к настоящему времени еще очень плохо изучен.
Защитным, приспособительным явлением у насекомых по отношению к некоторым болезнетворным микроорганизмам, в частности к некоторым бактериям, очевидно, служит фагоцитоз. Роль фагоцитов гемолимфы насекомых во многом подобна роли фагоцитов у высших животных. Отдельные фагоциты для разрушения инородных органических тел, поступивших в организм насекомых и имеющих большую, чем сами фагоциты, величину, собираются во временные скопления, образуют капсулу вокруг инородной частицы и коллективно ее разрушают, Постоянные скопления фагоцитов, или фагоцитоподобных клеток, имеются также в области спинного сосуда.
У некоторых насекомых выявлена секреция бактерицидных веществ. Так, аргентинский муравей fridomyrmex продуцирует иридомицин, близкий по сво- рис_ Лист насе,
ему химическому составу К не- КОМОЯДНОГО растения
петолактону, выделяемому гу- непентес (по Кер-
боцветными растениями Nepe- неру)
ta cataria L, (Паван и Пин-
четти, 1955). В секрете мандибулярных желез муравья Dendrolasius fuliginostis Latr. содержится бактерицидное вещество дендролазин (Паван, 1956). Силь- • ными бактерицидными свойствами обладает секрет сегментных желез личинок тополевого листоеда (Melasoma poputi L.) и личинок ивового листоеда {Phyllodecta vitetlina L.) (Взйн, 1943; Паван, 1953).
Наряду с фагоцитозом и секрецией биологически активных веществ, как средствами защиты от болезнетворных микроорганизмов, насекомые обладают, подобно высшим животным, гуморальной реакцией— способностью крови вырабатывать особые вещества — антитела, вступающие в химическую реакцию с проникшими в кровь чуждыми белками (антигенами) и их обеззараживающие.
Перитрофическая оболочка средней кишки, образующаяся у многих насекомых из выделений железистых клеток начальной части этого отдела пищеварительной системы, непроницаема для многих бактерий, поглощаемых вместе с пищей. Возникновение пе- ритрофической оболочки вызвано не только необходимостью предохранить тонкие эпителиальные стенки средней кишки от твердых частичек, о чем упоминалось в предыдущей главе, но и как приспособление, защищающее организм насекомого от некоторых заболеваний.
Все это касается, по-видимому, больше бактериальных, чем грибных заболеваний.
В отношении грибных заболеваний (как и бактериальных) у насекомых может существовать приспособительное поведение, препятствующее контакту с возбудителями заболеваний. Так, при изучении биологии кокцинеллид Brumus octosignatus Gebl. и Semiadalia undecimnoiata Schneid. мы обнаружили скопления жуков, зимующих в горах из гс|да в год в одних и тех же пунктах. Среди некоторых скоплений наблюдается массовая смертность жуков от заболевания, вызываемого грибом Tarichium sp. Оказалось, что такие пункты в следующем году не привлекают кокцинеллид. По-види- мому, ими руководит выработавшийся в качестве защитного приспособления отрицательный хемотаксис (различный запах больных и здоровых жуков в скоплениях отчетливо ощущается и человеком) (Яхонтов, 1950).
Представления о приспособительных особенностях растений, возникших для защиты от вредных насекомых, носят пока большей частью более или менее гипотетический характер. Предполагается, что некоторые анатомические черты растений, такие, как толщина кутикулы, опушонность, а также некоторые физиологические признаки (ядовитость и проч.), являются адаптациями по отношению к поедающим их животным, и особенно к насекомым. Хотя "известно, что едва ли не все виды растений подвергаются нападению насекомых, все же легко допустить, что названные признаки ограничивают число видов насекомых, способных к питанию за счет их, а те виды, которые поедают такие растения и вредят им, выработали ответные реакции, в частности, приспособили к ядовитым веществам характер своего метаболизма.
О том, что насекомые могут видоизменять характер метаболизма в зависимости от нищи, уже было сказано в предыдущей главе, а также в главе 5, где говорилось об образовании биологических рас и видов у насекомых. Еще в половине шестидесятых годов прошлого столетия Уолшем было замечено, что предпочтение кормового растения у многоядных видов насекомых может изменяться, а Гопкинсом было даже сформулировано особое правило выбора хозяина, согласно которому многоядным насекомым предпочитается всегда то растение, к которому оно приспособилось в предыдущих поколениях. Справедливость этого правила с начала настоящего столетия нашла подтверждение во многих специальных работах (см. главу 5).
Таким образом, в частности, произошли расы мухи Pegomyia kyosciami Panz. повреждающей свеклу, от мух, питавшихся раньше на белладонне и маревых (Chenopodiaceae) растениях (Камерон, 1914, 1916); так произошли и различные кормовые расы листоеда Phratora vitellinae L. (Шродер, 1903) и т. д. Поэтому в литературе подчеркивалось (Штейнер, 1925 и др.), что для правильного понимания поведения любой популяции насекомых важно учитывать поведение предыдущих генераций.
Регенерационная способность растений, более или менее быстрое восстановление объеденных насекомыми листьев, ветвей или корней, несомненно, является также защитной реакцией растений.
Некоторые авторы (Цвейгельт, 1928, 1931) склонны рассматривать даже образование полых галлов, внутри которых длительное время изолированы вредные насекомые, как реакцию защитного порядка, поскольку галлы ограничивают насекомых и препятствуют их расселению на весь период их изоляции.
В последнее время установлено, что у растений, подвергшихся нападению листогрызущих насекомых, энергия дыхания резко увеличивается, а это обеспечивает ускорение восстановительных процессов и интенсифицирует фотосинтез как оставшихся неповрежденных, так и,. вновь появляющихся листьев. При повреждениях листьев листогрызущими насекомыми деятельность ферментов растения приобретает большую направленность на вовлечение продуктов гидролиза в биосинтез и на связывание азота. . Срок деятельности листьев, оставшихся неповрежденными, удлиняется, и отток ассимилятов из листьев к растущим органам увеличивается. Все эти физиологические изменения являются несомненными признаками приспособительных реакций растений к повреждениям (Попов,
1959).
Происхождение приспособительных особенностей анатомического порядка в качестве защиты от нападения вредных насекомых кажется тем более вероятным, что подобные же приспособления по отношению к полезным насекомым уже давно выявлены и убедительно объяснены. Кроме того, подобные приспособления по отношению к растительноядным млекопитающим находили и экспериментальное подтверждение. Так, например, наличие колючек на некоторых растениях может препятствовать поеданию их скотом, и хотя колючки могут быть поняты так же, как средство к уменьшению потери влаги, почему такие растения особенно распространены именно в местностях с пустынным климатом, все же, когда Лютер Бербанк вывел искусственно кактусы без колючек, они4стали использоваться на корм скоту на пастбищах.
В качестве защитных от нападения бескрылых насекомых морфологических приспособлений истолковываются полосы направленных вниз щетинок, расположенных ниже узлов стебля Galeopsis L., или клейкие кольца на стеблях Lychnis viscaria, Silene viscosa, S. nutans (Крапелин, 1913).
Толщина кутикулы, очевидно, может иметь значение в отношении сосущих насекомых с коротким хоботком по аналогии с данными, полученными в последнее время для растениеядных клещей сем. Tetranychidae (Лебедев, Кособуцкий).
Советскими селекционерами выведены сорта подсолнечника, семена которых имеют углеродистый слой между пробковой тканью и склеренхимой, недоступный для челюстей гусениц подсолнечной огневки (Нотоео- soma nebulella Hb.). Такие сорта, получившие название «панцирных» или «молеустойчивых», совершенно не страдают от названного вредителя.
Для избежания нападения вредных насекомых мо-, жет иметь значение быстрота созревания растений. Так, например, на Яве жук из сем. Anthribidae на 75% повреждает семена Thephrosia Candida, но рано созревающие семена родственного растения Т. vogelli им не повреждаются. Конечно, выработалось ли это свойство в связи с жуком еще сомнительно, но самая мысль о подобных возможностях у растений логична.
Озимая совка (.Agrotis segetum Schiff.) нападает на хлопчатник только до времени одревеснения стеблей всходое. Характер онтогенеза злаков имеет во взаимоотношениях между ними и шведской мухой (Oscinosoma frit L.) решающее значение. Относительная устойчивость злаков определяется в основном скоростью начального развития и темпом увеличения конусов нарастания.
Установленный П. Г. Чесноковым (1956) факт варьирования устойчивости пшениц к повреждениям шведской мухи в зависимости от экологических условий (причем сорта, устойчивые на юге, обычно, оказываются неустойчивыми на севере и наоборот) получил новое освещение в работах Жуковского и Шапиро, оглашенных в 1960 г. на IV Всесоюзном энтомологическом съезде. Выяснилось, что устойчивый на юге сорт Ме- лянопус 69 медленно развивает конус нарастания на севере, на сорт Диамант большое влияние оказывают условия освещения. Но есть сорта, в равной степени устойчивые и неустойчивые в разных экол о го географических зонах. У устойчивых сортов Лютесценс 62, Це- зиум 111, например, конус нарастания и темп роста всходов происходят быстро в разных условиях освещения и температуры. У неустойчивого сорта Гор- диеформе 189, наоборот, начальные этапы онтогенеза замедлены как на севере, так и на юге.
В отношении влияния растительных ядов, ограничивающего возможность ’ нападения многих видов насекомых, указывалось, например, на атропин белладонны, на которой питаются лишь немногие виды. Очевидно, у таких насекомых выработался и специфический обмен веществ. Листоед Epithrix atropae Foudr., в частности, на других растениях не питается.
Свою специфическую фауну насекомых имеют крестоцветные растения, в соке которых содержатся глюко- зиды. Согласно Гагену (Фридерикс, 1932), древесина деревьев Tectona grandis и Sideroxylon не повреждается термитами из-за содержащихся в ней алкалоидов.
Обширные экспериментальные материалы и наблюдения имеются о защитном действии фитонцидов (в большинстве случаев это эфирные масла), которые в значительной мере относятся и к насекомым (Токин и др.). Здесь имеется полная аналогия в биологическом значении с антибиотическими веществами, выделяемыми многими низшими растениями. Согласно опытам Одесского сельскохозяйственного института смешанные посевы вики и растений, обладающих резко выраженными фитонцидными свойствами, — чеснока, лука, конопли и др. меньше страдали от акациевой огневки (Etiella zinckenella Тт.), чем чистые посевы вики. Меньше страдали от насекомых также посадки j капусты с помидорами и огурцами, бахчевых с коноплей и т. д. (Фуга, 1960).
В последнее время доказана связь устойчивости некоторых сортов растений по отношению к повреждениям сосущими насекомыми с осмотическим давлением клеточного сока этих растений (Соколов и Соколова, 1952; Яхонтов, 1956). Если осмотическое давление клеточного сока растения заметно превышает осмотическое давление полостной жидкости вредителя, растение почти не подвергается его нападению.
Насекомые должны также приспособляться и к защитным реакциям растений. С одной стороны, они избирают для повреждения менее защищенные растения, например, сорта с меньшим осмотическим давлением клеточного сока, с другой,— как уже упоминалось, у них может видоизменяться характер метаболизма. У некоторых насекомых известны даже специальные органы для освобождения организма от поглощаемых при питании на растениях вредных веществ. Таковы оеметерии гусениц некоторых видов Papilionidae, питающихся на растениях, содержащих эфирные масла и алкалоиды. Очевидно, такова же роль осметерий жуков из рода Malachius F. Оеметерии — вилообразно раздвоенные железы, выворачивающиеся под влиянием кровяного давления, у па-пилионид они выдвигаются между головой и первым сегментом груди сверху, у
Matachius F. по бокам переднегруди и среднегруди (рис. 68).
Осметериям часто приписывается отпугивающее воздействие на паразитов и хищников, но они при этом поглощают из крови и испаряют через свою поверхность во внешнюю среду ядовитые вещества. Вегенер (1923) подчеркивает, что у гусениц Papiiionidae, не питающихся ядовитыми растениями, эта железа слабо развита.
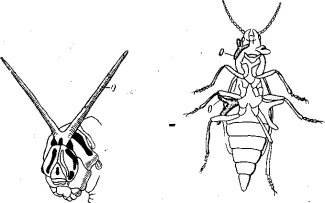
Рис. 68. Осметерии гусеницы Papilio macbaon L. (слева) и жука Malachias bipustulatus, L. (по Берлезе и Клеменцевичу):
о — осметерии
Хорошо известно защищающее от хищников действие обильного едкого секрета мешковидных кожных желез личинок жуков листоедов (Chrysomelidae), ¦ питающихся -на ивовых. Эти железы выделяют салициловую кислоту, которая вредна для организма, и поэтому их возникновение может рассматриваться так же как приспособление для питания на листьях ивовых деревьев (тополей, ив).
У гусениц листоверток (Tortricidae), питающихся на растениях, содержащих танин, в частности у зеленой дубовой листовертки (Tortrix viridana L.), в бокаловидных клетках средней кишки обнаружен танин в связанном состоянии, в виде кристаллов. Если же гусеницы эти кормятся на растениях, не содержащих в своем соке танина, такие кристаллы отсутствуют.
Очень велика полезная для растений роль насекомых в качестве переносчиков пыльцы для перекрестного опыления. Многие растения другим путем даже не могут опыляться и остаются бесплодными. Пыльца яблони, например, в силу своей влажности, даже при сильном ветре не переносится далее нескольких сантиметров и, как правило, не может попасть в другой цветок без помощи насекомого. Такие растения получили название энтомофильных. Полная и частичная энтомофилия стала возможной лишь благодаря взаимным приспособлениям со стороны растений и насекомых в процессе их эволюции. Так возникли нектаро- образование растений и аромат цветков, привлекающие насекомых-опылителей; так возникли строение цветков, удобное для опыления-насекомыми, и лепестки цветков,
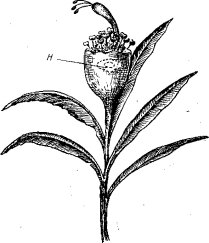
Рис. 69. Часть соцветия и прилегающих листьев Euphorbia pulcherrima Willd. (ориг.):
окрашенные в цвета, видимые насекомыми. Этим объясняется и почти полное отсутствие красной (без примеси фиолетового) окраски цветков в нашей флоре. Специальные исследования показали (Фриш, 1955, Чернышев, 1959 и др.), что все изученные насекомые, кроме бабочек рода Pie г is L., Epinep- hele jurtina L. и осы Peudovespa rufa L., почти или совершенно не воспринимают красный цвет.
н — нектарник
На рис. 69 изображена часть соцветия мексиканского растения Euphorbia pulcher- rima Willd., встречающегося в СССР только в оранжерейной и комнатной культуре. Цветки этого растения лишены лепестков, но листья близ соцветий имеют яркую красно-фиолетовую окраску. Комплекс мужских цветков и женский цветок заключены в общую оболочку, сбоку которой расположен нектарник. Здесь бросается в глаза специфическая приспособленность к энтомофильному опылению. Насекомое, привлеченное окраской верхних листьев и запахом нектара, чтобы достать этот нектар, вынуждено садиться на верх оболочки, из которой выступают цветки. Не менее выразительна приспособленность строения цветков у орхидных (Ог- chidaceae), губоцветных (Labiatae) (рис. 70), мотыльковых (Papiliona- сеае) и других энтомо- фильных растений.
1 — в разрезе. 2 — цветок с проникшим в него шмелем, 3 — положение пыльников до посещения насекомого, 4 — пыльники опустились под давлением шмеля на нижнюю губу венчика цветка; т — тычинка, п — пестик

Рис. 70. Цветок шалфея (по Тимирязеву):
У насекомых, в связи с их питанием нектаром и сбором пыльцы, возникло соответствующее устройство хоботка, а у специфических опылителей (группа пчелиных— Apodea) и специфическое строение задних ног; очевидно, в связи с опылительной деятельностью стоят и органы чувств (зрение, обоняние) и т. д.
Несмотря на очевидную взаимосвязанность эволюции насекомых и опыляемых ими цветковых растений, взаимные приспособления их все же относительны и несовершенны, поскольку в каждой группе, цветковых растений и в каждой группе опыляющих их насекомых существовали периоды, когда они имели независимое существование, т. е. или насекомые были связаны с другими растениями, или последние с другими группами насекомых. Так, например, пчелы рода Macropis Pz. встречаются лишь в местах, где есть вербейник (Lysi- machia), но ареал пчел значительно ;уже ареала вербейника. Появление и начальная эволюция вербейника много древнее, чем у пчел названного рода (Попов, 1958).
Наиболее хорошо изучена опылительная деятель-
Н'оеть домашней пчелы (Apis mellifera L.) и по этому вопросу имеется обширная пчеловодческая литература, В настоящее время крупные садоводческие хозяйства организуют специальные опылительные пасеки, поскольку опыт показал, что урожай плодовых деревьев при этом сильно возрастает. Работа автора по изучению роли диких опылителей яблони в Узбекистане {Яхонтов, 1946) показала, что без искусственного пчелоопы- ления могут с успехом использоваться лишь небольшие садовые участки, с близко находящимися гнездами диких пчел или наличием ряда других, второстепенных в смысле опыления насекомых, способных переносить пыльцу яблони с цветка на цветок. Даже такие цветковые растения, как, например, хлопчатник, которые способны к самоопылению, в значительной мере посещаются опылителями, и роль последних в опылении весьма значительна (Яхонтов, 1936, 1937).
Классическим примером опылительной роли насекомых в жизни растений является завоз красного клевера в Австралию, где не удавалось получить семян этого растения до тех пор, пока в Австралию не были завезены шмели (Bombidae). Из этого пример а видно также, что взаимное приспособление растений и насекомых может предусматривать определенные их виды или группы. Одни цветки (садовая ночная красавица) открываются только ночью и опыляются некоторыми видами насекомых, активных в ночное время, другие ¦ (одуванчики) на ночь, наоборот, закрываются и нектар выделяют только . днем. У растений, опыляемых бабочками (Lepidoptera)г нектар находится обычно в глубоких чашечках цветков и не доступен для большинства других насекомых. Растения из семейства губоцветных и бобовых приспособлены преимущественно к опылению перепончатокрылыми насекомыми (Нугпе- noptera).
Чрезвычайно своеобразные взаимоотношения известны у юкки (Jucca filamentosa) и опыляющей ее цветки юкковой моли (Pronuba fuccasella Riley.), а также у инжира и его специфического опылителя — халь- цида блаетофаги (Blastophaga psenes L.). Для обеспечения перекрестного опыления растение теряет даже' часть своих генеративных образований.
Самка юкковой моли при помощи щупалец соби-
рает с тычинок юкки влажную пыльцу, затем откладывает внутрь пестика другого цветка яйцо и укладывает на рыльце пестика принесенную ею пыльцу. Некоторые из образующихся, завязей в дальнейшем уничтожаются гусеницей моли, но без моли юкка семян не производит. Подобные же отношения, хотя и менее детально изученные, отмечены и у других видов юкк юго-запада США и Мексики с другими видами юкковых молей того же рода. Каждый вид моли способен опылять только определенный вид юкки. Юкки цветут не каждый год, поэтому их опылители способны к очень длительной диапаузе (в стадии куколки).
Дикий инжир, а также некоторые сорта культурного инжира, не способны к самоопылению и без бластофаги семян не образуют. У них различают две категории деревьев — каприфиги и фиги. Первые не плодоносят и развивают мужские и короткопестиковые женские цветки; цветки фиг длиннопестиковые. Бластофага развивается вйутри коротких пестиков на каприфйгах, превращая их в галлы. Бескрылые самцы в полом цветоложе оплодотворяют крылатых самок, которые, покидая цветоложе, бывают обильно опудрены пыльцой инжира. Затем они проникают к цветкам фиг и обеспечивают их опыление, но яйца в длиннопестиковые цветки из-за короткого яйцеклада они отложить не могут. Осенние соцветия инжира имеются только на каприфи- гах, в них происходит зимовка бластофати. Весной соцветия развиваются преимущественно на каприфитах, где откладываются яйца перезимовавшим поколением опылителя, летом же преобладают цветки на фигах.
Имеется много и таких видов растений, цветки которых доступны для опыления весьма разнообразными насекомыми. Опыление может произойти конечно лишь в том случае, если насекомое будет переносить на теле пыльцу в другой цветок того же вида растения. Некоторые насекомые, однако, будучи привлечены нектаром и напитавшись, в другие цветки не стремятся; при таких условиях насекомое приносит растению уже не пользу, а вред. .
Насекомых, не способных к опылению посещаемых ими цветков (большинство муравьев — Formicidae или некоторые жуки, например Bruchidae, Dermestidae и ДР-), называют д и с т р опными. Насекомых, не имеющих специфического поведения, необходимого для опыления цветков, но иногда переносящих на себе пыльцу из цветка в цветок, например, многих мух (Diptera) или ос (Vespidae), относят к аллотропным. Специфических опылителей цветов, таких, как бабочки бражники (Sphingidae) или большинство пчелиных насекомых (Apodea), называют евтропными.
Естественно, что евтропные насекомые имеют для растений наибольшее значение в качестве опылителей. Адаптивные признаки у растений могут все же возникать и по отношению к аллотропным насекомым — такие цветки имеют нектар и пыльцу в верхних их частях, например Euphorbia. А у некоторых растений (Arum) создались цветки-ловушки, задерживающие аллотропных насекомых (мух) до тех пор, пока опыление не будет обеспечено.
Конечно, по отношению к дистропным насекомым у растений могут образовываться защитные приспособления. К ним относят, например, густые сплетения волосков в венчиках цветков растений родов Ме- nyanthes и Thymus и кольца из волосков, задерживающие муравьев у Lamium и Stachys. К защите нектара цветков от некоторых дистропных насекомых в большой мере могут относиться и вышеупомянутые клеевые кольца на стеблях растений.
На примере евгропных и отчасти аллотропных насекомых мы видим взаимно полезное сожительство их в биоценозах с растениями. Еще более тесные симбиотические отношения отмечены между некоторыми насекомыми и населяющими их кишечник дрожжевыми грибами и бактериями (Вернер, 1927; Хиц, 1927 и др.).
Такие грибы и бактерии живут и размножаются за счет пищевых масс, а иногда и за счет клеточного сока насекомых, не принося им даже в последнем случае сколько-нибудь ощутительного вреда. Эти грибы и бактерии выделяют ферменты, необходимые для усвоения некоторых частей пищи насекомых (например, крови высших животных и человека у кровососущих видов, клетчатки, сахаров). Подобные симбионты выявлены у вшей (Parasita), комаров (Culicidae), некоторых тлей (Aphidodea), кокцид (Coccodea), личинок бронзовки Poiosia meiallica Hbst. и др. У черного таракана (Blatta orientalis L.) имеются скопления симбионтов и в жировом теле. Эти скопления получили название м и- ц е т о м.
Полезные для растений и насекомых связи устанавливаются тогда, когда они обеспечивают более широкое расселение видов. В прошлой главе уже говорилось, что некоторые виды грибов даже «культивируются» термитами (Isoptera). Семена и споры растений несомненно расселяют многие насекомые, особенно муравьи (Formicidae). Сеноед Lachesilla un-sui Danks. распространяет споры гриба Nigrospora gossypii Jacz (Цой Ун-су, 1951); жуки заболонннки (Scolitinae) разносят споры и мицелий гриба Graphium ulmi Schw.—¦ возбудителя голландской болезни вязов и т. д. Разнос семян и спор на поверхности тела животных носит название эктоз'оохории. Разнос синантропы ым и насекомыми—домашней мухой (Musca domestica L.) и другими — болезнетворных бактерий хорошо изучен в медицинской энтомологии.
С другой стороны, и растения помогают расселению : насекомых. В опавшей осенью листве деревьев залегают ’¦ на зимовку многие насекомые. Уносимые ветром листья могут переносить на себе и находящихся на них насекомых. Так, в системе карантинных мероприятий^ препятствующих расселению червеца комстока (Pseudococcus comstocki Kuw.) в новые места в Узбекистане, Таджикистане и Грузии, одно из важных мест занимает уничтожение опавших с деревьев листьев.
Распространяются с семенами, например, многие : жуки зерновки (Bruchidae), в том числе серьезные вредители сельского хозяйства — гороховая (Bruchus pi- sorum L.) и фасолевая (Acanthoscelides obtectus Say.) зерновки, некоторые долгоносики (Curculionidae), в том числе амбарные вредители — рисовый 1 (Calandra огу- zae L.) и амбарный (С. granaria L.) долгоносики. Расселяются в летучках семян тау-сагыза закокониро- вавшиеся личинки таусагызной галлицы (Perrisia tau- saghyzae Dombr.) в Казахстане (Правдин) и многие другие насекомые.
Подробно говорить о роли растений в качестве убежища (укрытия) для насекомых от хищников и неблагоприятных погодных условий, видимо, нет необходимости. О -зимовке насекомых под опавшей листвой деревьев, где насекомые лучше сохраняются от морозов, говорилось раньше. Насекомые могут зимовать и в других растительных остатках (соломе и т. д.), под чешуйками коры деревьев и т. д. Во время дождя многие насекомые находятся с нижней стороны листьев, где они могут пребывать и во время благоприятной погоды, будучи в состоянии покоя. Это, очевидно, помотает им избежать нападения летающих над растениями паразитов и хищников. Во время жаркого периода суток, с нижней стороны листьев хлопчатника, например, можно видеть укрывшимися многих насекомых, не имеющих пищевых связей с этим растением. При повышенной температуре личинки азиатской саранчи (Locusta migrator Іа L.) и мароккской кобылки (Dociostaurus marocca- nus Thnb.) забираются в тень растений и т. д.
Связи насекомых с растениями, таким образом, очень многообразны. Эти связи часто определяют-группировки видов и распределение популяций в биотопах (см. главы 3 и 5). Численность насекомых в биоценозах в значительной мере определяется также этими связями, равно как и численность растений, в свою очередь, может в большой степени определяться вредной и полезной для них деятельностью насекомых. В общих чертах обилие пригодных для питания того иди иного насекомого растений является фактом, благоприятным для роста численности этого насекомого и, наоборот, обилие насекомого фитофага может угнетать численность растений. Эти противоречивые отношения между растениями и насекомыми фитофагами — одна из причин, обусловливающих колебания численности как тех, так и других.
Во вторичных биотопах иногда может даже происходить почти полное искоренение отдельных видов растений насекомыми на больших площадях. По высказыванию Голлоуэй \ уже проводилась успешная борьба с сорняком Hypericum perforatum L. при помощи питающихся на нем насекомых во всех районах Калифорнии, а, по утверждению Смиса \ специально завезенные насекомые фитофаги подавляют размножение сорняка Eupatorium adenophorum Spreng. на больших территориях на Гавайских островах; те же результаты были получены на острове Гонолулу (Фаллауэй) [11] при помощи насекомых в отношении портящего пастбища кактуса Opuncia megacantha.
Еще по теме ОСОБЕННОСТИ ВЛИЯНИЯ БИОТИЧЕСКИХ ФАКТОРОВ. ЭКОЛОГИЧЕСКИЕ СВЯЗИ НАСЕКОМЫХ С РАСТЕНИЯМИ:
- Глава И ЭКОЛОГИЧЕСКИЕ СВЯЗИ НАСЕКОМЫХ МЕЖДУ СОБОЙ И С ДРУГИМИ животными
- ВЛИЯНИЕ АНТРОПОГЕННЫХ ФАКТОРОВ НА НАСЕКОМЫХ
- ПИЩА КАК ФАКТОР СРЕДЫ И ЕЕ ВЛИЯНИЕ НА НАСЕКОМЫХ
- Глава 8 ВЛИЯНИЕ НА НАСЕКОМЫХ СВЕТА И ДРУГИХ АБИОТИЧЕСКИХ ФАКТОРОВ СРЕДЫ
- Глава III Растение и среда. Значение экологических факторов в жизни бромелиевых
- Биотические и антропогенные факторы
- Биотический потенциал насекомых
- Нарушение биотического равновесия под влиянием деятельности человека
- Экологические факторы
- 1.2. Экология и экологические факторы
- СРЕДА И ЭКОЛОГИЧЕСКИЕ ФАКТОРЫ
- Г лав а 6 ВЛИЯНИЕ ТЕМПЕРАТУРЫ НА НАСЕКОМЫХ
- Г лава 4 ЭКОЛОГИЧЕСКИЕ НИШИ И ЖИЗНЕННЫЕ ФОРМЫ НАСЕКОМЫХ
- Влияние влажности на насекомых
- ПОЧВЕННЫЕ (ЭДАФИЧЕСКИЕ) ФАКТОРЫ СРЕДЫ НАСЕКОМЫХ
- ВЛИЯНИЕ СООБИТАТЕЛЕЙ НА ПОЛОЖЕНИЕ ЭКОЛОГИЧЕСКОГО ОПТИМУМА
- Глава 7 ВЛИЯНИЕ НА НАСЕКОМЫХ ВЛАЖНОСТИ И ОСАДКОВ
- Влияние на насекомых низких и высоких температур
- ГЛАВА 5. ЭКОЛОГИЧЕСКАЯ ЭВОЛЮЦИЯ НАСЕКОМЫХ
- Влияние температуры на поведение насекомых