ПОСЛЕСЕМЕННОЕ РАЗВИТИЕ И МЕТАМОРФОЗ У THUNIA MARSCHALLIANA
Начало изучению особенностей образа жизни и развития орхидей было положено работами Бернара (Bernard, 1899, 1904, 1909), который установил облигатность симбиотического микотрофного развития гнездовки (Neottia nidus-avis).
В дальнейшем обогащению наших знанийо развитии и образе жизни орхидей во многом способствовали работы таких интересных исследователей, как Бургефа (Burgeff, 1909,f 1932, 1943, 1954), Pay (Rauh, 1937), Фермейлена (Vermeulen, 1947), В. А. Поддубной-Арнольди (1959а, 19596, 1960, 1964а, 19646), Вейре (Veyret, 1965).
К сожалению, исследования, посвященные изучению послесеменного развития орхидей, касались главным образом внешней, морфологической стороны развития, не было достаточно полной и точной картины происходящих морфогенетических преобразований.
Мы изучали послесеменное развитие у Thunia marshalliana (Терёхин, 19686; Терёхин, Никитичева, 1968). Хотя это растение относится к одной из наименее специализированных групп в сем. Orchidaceae, тем не менее ему присущи все основные черты метаморфоза, характерные, по-видимому, для большинства представителей этих интересных паразитных цветковых.
Зрелое семя Thunia содержит зародыш, в котором можно довольно отчетливо различить зачаток семядоли. Для зародыша характерно полное отсутствие признаков радикулярной организации (рис. 33, I). Послесеменное развитие Thunia изучалось нами на искусственной среде.
Начало прорастания семени Thunia характеризуется диффузным разрастанием тела зародыша-проростка (протокорма, протосомы), происходящим главным образом за счет роста клеток (в дальнейшем в морфогенетических процессах начинают играть все большую роль неориентированные и ориентированные клеточные деления). Зачаток первой семядоли располагается на зародыше не терминально, а несколько смещен к одной из его боковых сторон (рис. 33, 2); он обнаруживает трубчатый характер развития и образует вокруг терминальной области зародыша-протосомы своеобразное «влагалище» (рис.
'33, 2—5, 34). В основании влагалища закладывается примордий второй семядоли, развитие которой также имеет трубчатый характер. Тело проростка-протосомы продолжает в это время диффузно разрастаться, а примерно в середине его терминальной области, в основании морфогенетически активной зоны, может быть замечена дифференциация прокамбиального тяжа. В основании второй семядоли возникает апекс побега, который в дальнейшем последовательно отчленяет зачатки листьев (рис. 33, 6—22). Отмеченное выше явление дифференциаиии второй семядоли из тканей проэмбрио — факт, требующий, по-видимому, подтверждения примерами послесеменного развития у других, видов орхидей.В латеральных и базальных областях протосомы (протокорма) образуются эпидермальные волоски, которые в естественных условиях служат, по-видимому, поставщиками воды и минеральных веществ; через эпидермальные волоски часто также происходит внедрение гиф микоризного гриба в ткани цветкового растения (Harley, 1959; Schmucker, 1959а). Бернар (Bernard, 1909) наблюдал в естественных условиях развитие очень сходной с Thunia по уровню специализации орхидеи Bletilla hyacinthina- Он отметил выделение в теле зародыша-протосомы особых гаусториальных зон, в клетках которых поселяется микоризный гриб и где происходит его «усвоение» цветковым симбионтом.
Через определенное время после начала функционирования апекса побега у Thunia, как и у большинства изученных к настоящему времени орхидей, наблюдается образование и развитие первых адвентивных корней, возникающих на стебле обычно выше основания семядолей (Bernard, 1909; Burgeff, 1936; Поддубная-Арнольди, Селезнева, 1957; Поддубная- Арнольди, 19646; ,Терёхин, Никитичева, 1968, и др.). Эти корни также атакуются микоризным грибом.
В процессе развития адвентивной корневой системы протокорм (зародыш-протосома) отмирает и структура орхидеи к моменту цветения в основных своих чертах принимает биполярную организацию типичного автотрофного цветкового растения. Описанный способ морфогенетических преобразований, как мы увидим дальше, претерпел в процессе эволюции Орхидных значительные изменения (например, в направлении сохранения зародыша-протосомы в течение всего онтогенеза растения у Corallorhiza или Epipogium).
Рис. 33. Послесеменное развитие Thunia marschalliana (сем. Orchidaceae).
л—11 — последовательность стадий. C1 — первая семядоля; с2 — вторая семядоля; л%—лъ — первый и последующие листья; ап — апекс побега; к — корень; пх — первичный протокорм; п2—п3 — вторичный протокорм.
Развитие спорофита Thunia — типичный пример развития с метаморфозом, характерный для представителей четвертой формы паразитизма. Краткая схема метаморфизованного развития Thunia выглядит следующим образом. Униполярное развитие проростка из терминальной области зародыша после внедрения гиф микоризного гриба в клетки его базальной области. Становление биполярной организации (типа побег—микоризный корень) из вторичных меристематических очагов.
Рис. 33 (продолжение).
3. Отмирание выполнивших свою функцию тканей протокорма.
Здесь, так же как и в других типах метаморфизованного развития, характер структурных преобразований спорофита соответствует функциональным потребностям нового образа жизни. Резервация пищевых веществ и связанное с этим каллюсообразное разрастание проростка (как и у некоторых Заразиховых, Баланофоровых и т.д.) является базой для определенного типа полового и вегетативного воспроизведения, когда одновременно в теле проростка закладывается не один, а несколько ини- циалей генеративных побегов (своеобразная полиэмбриония, по В. А. Под- дубной-Арнольди, 19646).
Рис. 34. Серии поперечных срезов? через протокорм Thunia marschal- Iiana (сем. Orchidaceae).
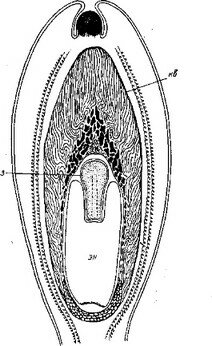 А — заложение второй семядоли в пазухе первой; Б — развитие семядолей; В — заложение апекса побега в пазухе второй семядоли. ct — первая семядоля; с2 — вторая семядоля; ап — апекс побега.
А — заложение второй семядоли в пазухе первой; Б — развитие семядолей; В — заложение апекса побега в пазухе второй семядоли. ct — первая семядоля; с2 — вторая семядоля; ап — апекс побега.
На рис. 35 схематично изображены основные способы по- слесеменного развития зародыша у представителей каждой из четырех основных форм чужеядности. В отличие от автотрофных и слабоспециализированных паразитных растений (рис. 35,1,2), где у проростка одинаково активны апикальный и базальный полюсы, у Orobanche и Pyrola (рис. 35, 3, 5) развивает
ся в послесеменном периоде только базальный полюс зародыша, тогда как апикальный отмирает; у Cuscuta1 Neottia и Thunia (рис. 35, 4—6), напротив, при прорастании развивается морфогенетически только апикальный полюс зародыша.
Как мы могли видеть на примере Thunia1 даже наименее специализированные виды орхидей имеют метаморфизованный способ
развития спорофита. Основной особенностью метаморфизованного разви
тия спорофита у Орхидных является то, что здесь вторичное становление биполярной организации растения происходит не вследствие развития адвентивных (по своему эволюционному происхождению) побегов, но вследствие появления адвентивных корней.
Еще по теме ПОСЛЕСЕМЕННОЕ РАЗВИТИЕ И МЕТАМОРФОЗ У THUNIA MARSCHALLIANA:
- ПОСЛЕСЕМЕННОЕ РАЗВИТИЕ И МЕТАМОРФОЗ У OROBAN С HE CREN AT А И CY NOMORIU M SONGARICUM
- ПОСЛЕСЕМЕННОЕ РАЗВИТИЕ И МЕТАМОРФОЗ У ПРЕДСТАВИТЕЛЕЙ СЕМ. PYROLACEAE
- ПОСЛЕСЕМЕННОЕ РАЗВИТИЕ, МЕТАМОРФОЗ И ЭВОЛЮЦИЯ ПРОТОСОМЫ У CUSCUTA
- МЕТАМОРФОЗ
- МЕТАМОРФОЗ, РЕДУКЦИЯ И НЕОТЕНИЯ И СИСТЕМЫ МОДУСОВ ЭВОЛЮЦИОННЫХ СТРУКТУРНЫХ ПРЕОБРАЗОВАНИЙ
- ПРОДОЛЖИТЕЛЬНОСТЬ РАЗВИТИЯ
- 18.7. ЦИКЛ РАЗВИТИЯ ПАРАЗИТОВ И ОРГАНИЗМ ХОЗЯИНА
- Канализация развития
- 6. 4. 7. Развитие биосферы
- Эмбриональное развитие
- ЭМБРИОНАЛЬНОЕ РАЗВИТИЕ
- Влияние температуры на развитие насекомых.
- Динамика развития гибридов кукурузы
- 8.2.7. Генетический контроль развития
- 4.2. Амплификация в процессе развития
- 9.2. КЛАССИФИКАЦИЯ ВРОЖДЕННЫХ ПОРОКОВ РАЗВИТИЯ
- Мозаичная теория развития
- 13.3.4. Атавистические пороки развития
- РАЗВИТИЕ ФИТОВИРУСОЛОГИИ
- РАЗМНОЖЕНИЕ И РАЗВИТИЕ РЫБ