ПОСЛЕСЕМЕННОЕ РАЗВИТИЕ И МЕТАМОРФОЗ У ПРЕДСТАВИТЕЛЕЙ СЕМ. PYROLACEAE
Начало изучению особенностей послесеменного развития высокоспециализированных микотрофных растений на примере представителей сем. Pyrolaeeae было положено Беленовским (Velenovsky, 1892, 1905, 1907).
Беленовский на основании полевых наблюдений сформулировал гипотезу, согласно которой в жизненном цикле спорофита некоторых грушанок можно выделить две генерации: подземную генерацию, названную им «прок аул омом», живущую вполне паразитически за счет микоризного гриба, и надземную генерацию — облиственные, зеленые генеративные побеги, которые существуют за счет фотосинтеза и биопродуктов микоризы. Прокаулом представляет собой, по мнению Беленовского, нечто среднее между стеблем и корнем и по своей структурной организации сходен с подземными образованиями у представителей семейств Orobanehaeeae и Balanophoraeeae.Как мы уже видели, зародЬшш в зрелых семенах Pyrola, Moneses9 Hypopitys представлены немногочисленными овальными образованиями, в которых можно различить эмбриодерму, базальную меристему и основную ткань (Pyrola9 Moneses), либо эти зародыши состоят из 2—5 гистологически недифференцированных клеток (Hypopitys и др.). При прорастании семени грушанок и вертляниц происходит униполярное развитие проростка из базальной области зародыша. Апикальная область зародыша остается в семени, где и отмирает (рис. 26, I—5), выполнив гаусториальную функцию в эндосперме (Christoph, 1921; Rauh, 1937; Francke, 1934— 1935; Ltick, 1940, 1941; Lihnell, 1942). Проросток-протосома развивается униполярно и ветвится (рис. 26, 4—6). Каллюсообразного разрастания (клубенька) не образуется. Линель (Lihnell, 1942) отметил, что вегетативное развитие проростков на искусственной среде до образования генеративных побегов может продолжаться по меньшей мере в течение трех с половиной лет. Становление биполярной организации происходит при переходе к половому воспроизведению за счет образования адвентивных побегов.
Таким образом, и здесь мы обнаруживаем типичную, хотя и своеобразную картину метаморфоза — смену униполярного биполярным способом развития и организации, некроз тканей, выполнивших свою функцию, становление структур биполярной организации из вторичных меристематических очагов (Терёхин, 19686; Терёхин, Никитичева, 1968).
При изучении Грушанковых нельзя не обратить внимания на одну весьма характерную для этой группы растений особенность развития.
 alt="" />
alt="" /> 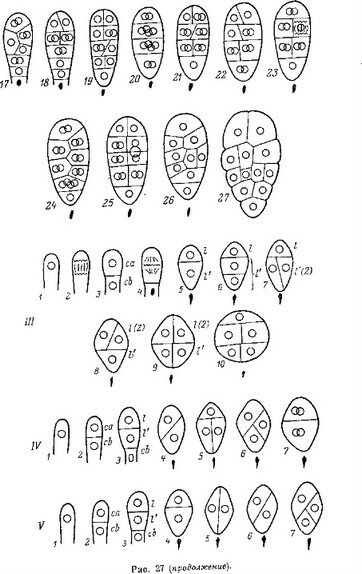
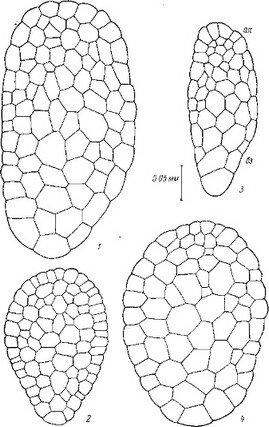
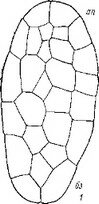 Рис. 26. Послесеменное развитие некоторых представителей сем. Pyrolaceae.
Рис. 26. Послесеменное развитие некоторых представителей сем. Pyrolaceae.
I — pyrola гotundifolia\ 2 — Orthilia secunda\ 3 — Moneses uniflora (по: Luck, 1940); 4—6 — P. ro- tundifolia (по: Lihnell, 1942). an — апикальный полюс зародыша; бз — базальный полюс зародыша;
эн — эндосперм.
Вскоре после начала прорастания (для разных видов с разными уровнями специализации этот период различен) развитие проростка приостанавливается и в естественных условиях продолжается далее только в случае контакта с подходящим грибным мицелием (Francke, 1934—1935; Liick, 1940, 1941; Lihnell, 1942, и др.). Гриб внедряется в ткани, расположенные под морфогенетически активной верхушечной зоной проростка. У более специализированных видов (Hypopitys) приостановка развития происходит онтогенетически раньше.
Интересно отметить, что заложение и развитие органов полового воспроизведения у некоторых грушанок (Pyrola, Orthilia) происходит в два этапа (Терёхин, 1962г) Сначала образуются многолетние корневища, а уже в них в каждый вегетационный сезон закладывается один или несколько генеративных побегов.
ЭВОЛЮЦИЯ ПРОТОСОМЫ
Остановимся на некоторых основных тенденциях в эволюции подземных стадий послесеменного развития спорофита.
Сем. Pyrolaceae. Специализация послесеменной стадии развития спорофита в сем. Грушанковых идет в направлении все более глубокого преобразования корневых структур в структуры протосомы. Здесь можно выделить два этапа специализации. Представителем первого может служить Pyrola rotundifolia. Проросток-протосома этого растения развивается из базальной области зародыша и до начала контакта с микоризным грибом проходит две стадии асимбиотического развития: конусовидную и ци
линдрическую. К этому времени запасы питательных веществ в эндосперме иссякают и развитие протосомы приостанавливается до контакта с микоризным грибом, который активно проникает в ткани базальной области проростка (Christoph, 1921; Liick, 1941).
Беленовский (Velenovsky, 1907) предполагал, что, в отличие от более высокоспециализированных Moneses и Hypopitys, проросток P. rotundb- folia развивается после контакта со своим паразитным грибом биполярно, т. е. обычным для автотрофного растения образом. Однако Pay (Rauh, 1937) и Люк (Liick, 1940)*высказали сомнение относительно того, что спо- соб прорастания Pyrola может значительно отличаться от такового у Mo- neses и Hypopitys. Вскоре Линель (Lihnell, 1942) доказал экспериментами по прорастанию семян Pyrola и Hypopitys справедливость этих сомнений: протосома P. rotundifolia в течение 3.5 лет развивалась униполярно.
В результате длительных полевых наблюдений за развитием P. rotundifolia нам удалось обнаружить, что у этого растения протосома дает начало корневищу, и уже на этом последнем образуются генеративные побеги. Таким образом, здесь мы обнаруживаем тот же способ развития, что и у Moneses и Hypopitys1 но между протосомой и генеративным побегом здесь как бы вклинивается дополнительная структура — корневище. Присутствие корневища, органа, характерного для автотрофных растений, свидетельствует о более низком уровне специализации спорофита Pyrola в сравнении со спорофитами Moneses и Hypopitys.
У P. rotundifolia протосома, в отличие от протосом Moneses и Hypopitys1 после развития корневища отмирает, и в этом мы видим еще одно различие в уровнях специализации спорофитов данных растений в направлении паразитизма.В отдичие от Pyrola1 у Hypopitys асимбиотическое прорастание затруднено и асимбиотический проросток протосомы развивается самостоятельно только до конусовидной стадии (Francke, 1934—1935; Liick, 1941; Lihnell, 1942). У Moneses Uniflora1 Hypopitys monotropa, Monotropa uniflora и, по- видимому, у Pterospora andromeda, Sarcodes sanguinea и других вертляниц протосома существует очень долго, вероятно в течение всего онтогенеза. Вегетативное размножение осуществляется здесь за счет образования новых генеративных побегов на ветвящейся протосоме (С3-состояние протосомы).
Для всех Грушанковых в послесеменном развитии характерна стадия поиска питающего субстрата, выраженная в состоянии ожидания атаки микоризного гриба. Эта стадия имеет вид типичного Л-состояния униполярно развивающейся в этот период протосомы. 5-состояние протосомы у Вертляницевых нет в связи с характером ее размножения путем ветвления первичной Л-протосомы. Очевидно, формула послесеменного развития в этом семействе может быть представлена для всех изученных его представителей сочетанием символов ЛС3-состояния протосомы (см. табл. 2).
В сем. Грушанковых мы обнаруживаем те же формы эволюционного преобразования послесеменной стадии спорофита, что и в рассмотренных выше группах ксенопаразитов. Как у Норичниковых и Заразиховых, в эволюции послесеменной стадии развития спорофита Грушанковых мы обнаруживаем уже закончившийся в сущности процесс субституции основного побега адвентивными структурами на основе образования новой для этих растений структуры — протосомы.
Сем. Gentianaceae. Фигдор (Figdor, 1897) нашел в природе молодые проростки Cotylanthera tenuis. Автор отмечает, что у зародыша прорастает только его базальный (корневой) полюс, в то время как его терминальная (неактивная) область остается в семенной оболочке.
Анатомия зрелого семени также вполне определенно свидетельствует об активности базальной области проэмбрио в ранние периоды прорастания. Опираясь на данные Фигдора, можно думать, что у паразитных видов семейства также представлены А- и С3-состояния протосомы.Сем. Burmanniaceae и Thismiaceae. Строение зрелого семени Бурманние- вых показывает, что протосома этих растений развивается из базального полюса проэмбрио. Джонкер (Jonker, 1938) сообщил, что представители секции Foliosa рода Burmannia имеют многолетние корневища, что сближает их по уровню специализации с Pyrola (Pyrolaceae). В то же время остальные представители семейства обладают типичными протосомами (подобными таковым Moneses или Hypopitys) на протяжении всего их жизненного цикла (Johow, 1889; Ernst, Bernard, 1912; Goebel, Suesenguth, 1924; Rauh, 1937; Schmucker, 1959a, и др.). Известно, что для Burmannia сое- Iestis характерен прокаулом с главным телом и огрибленными ответвлениями. В. Candida (так же как и виды Thismia) имеет толстые короткие прокауломы, из которых эндогенно возникают генеративные побеги.
Для Thismia Iutzenburgii и Burmannia championii характерно присутствие 5-состояния протосомы (Johow, 1889; Rauh, 1937; Schmucker, 1959а, и др.). Однако у других видов 5-состояние протосомы, очевидно, не выражено, а встречаются только два состояния — А и С3.
Сем. Triuridaceae. Строение зрелого семени и уровень дифференциации зародышей свидетельствуют о том, что проросток Sciaphila развивается из базального полюса проэмбрио.
Таким образом, для всех изученных представителей группы растений с третьей формой паразитизма характерна высокая специализация способов послесеменного развития и строения проростков-протосом. Сходное в общих чертах направление этой специализации указывает еще раз на общность факторов их эволюции.
Еще по теме ПОСЛЕСЕМЕННОЕ РАЗВИТИЕ И МЕТАМОРФОЗ У ПРЕДСТАВИТЕЛЕЙ СЕМ. PYROLACEAE:
- ПОСЛЕСЕМЕННОЕ РАЗВИТИЕ И МЕТАМОРФОЗ У THUNIA MARSCHALLIANA
- ПОСЛЕСЕМЕННОЕ РАЗВИТИЕ И МЕТАМОРФОЗ У OROBAN С HE CREN AT А И CY NOMORIU M SONGARICUM
- ПОСЛЕСЕМЕННОЕ РАЗВИТИЕ, МЕТАМОРФОЗ И ЭВОЛЮЦИЯ ПРОТОСОМЫ У CUSCUTA
- ЭНДОСПЕРМ РАСТЕНИЙ СЕМ. CUSCUTACEAE
- ЭВОЛЮЦИОННЫЕ ПРЕОБРАЗОВАНИЯ У РАСТЕНИЙ С ТРЕТЬЕЙ ФОРМОЙ ПАРАЗИТИЗМА (ФОРМА «PYROLACEAE»)
- МЕТАМОРФОЗ
- МЕТАМОРФОЗ, РЕДУКЦИЯ И НЕОТЕНИЯ И СИСТЕМЫ МОДУСОВ ЭВОЛЮЦИОННЫХ СТРУКТУРНЫХ ПРЕОБРАЗОВАНИЙ
- ОБЩАЯ ХАРАКТЕРИСТИКА ПРЕДСТАВИТЕЛЕЙ СЕМЕЙСТВА REOVIRIDAE
- ОБЩАЯ ХАРАКТЕРИСТИКА ПРЕДСТАВИТЕЛЕЙ СЕМЕЙСТВА Circoviridae
- ОБЩАЯ ХАРАКТЕРИСТИКА ПРЕДСТАВИТЕЛЕЙ СЕМЕЙСТВА FLAVIVIRIDAE
- ОБЩАЯ ХАРАКТЕРИСТИКА ПРЕДСТАВИТЕЛЕЙ СЕМЕЙСТВА TOGAVIRIDAE
- ОБЩАЯ ХАРАКТЕРИСТИКА ПРЕДСТАВИТЕЛЕЙ СЕМЕЙСТВА CALICrVTRIDAE
- ОБЩАЯ ХАРАКТЕРИСТИКА ПРЕДСТАВИТЕЛЕЙ CORONAVIRIDAE
- ОБЩАЯ ХАРАКТЕРИСТИКА ПРЕДСТАВИТЕЛЕЙ СЕМЕЙСТВА BIRNAVIRIDAE
- ОБЩАЯ ХАРАКТЕРИСТИКА ПРЕДСТАВИТЕЛЕЙ СЕМЕЙСТВА PARAMYXOVIRIDAE
- ОБЩАЯ ХАРАКТЕРИСТИКА ПРЕДСТАВИТЕЛЕЙ СЕМЕЙСТВА PICORNAVnODAE
- ОБЩАЯ ХАРАКТЕРИСТИКА ПРЕДСТАВИТЕЛЕЙ СЕМЕЙСТВА ARTERIVIRIDAE
- ОБЩАЯ ХАРАКТЕРИСТИКА ПРЕДСТАВИТЕЛЕЙ СЕМЕЙСТВА ADENOVIRIDAE