ПОСЛЕСЕМЕННОЕ РАЗВИТИЕ, МЕТАМОРФОЗ И ЭВОЛЮЦИЯ ПРОТОСОМЫ У CUSCUTA
Cuscuta давно является объектом пристального внимания исследователей как злостный вредитель многих сельскохозяйственных культур (Koch, 1877, 1880а; Macpherson, 1921; Fedortschuk, 1931; Marloth, 1932; Tiagi, 1951а; Johri, Tiagi, 1952; Пилюгин, 1953; Мещеряков, 1954; Еленев, 1956; Котт, 1959; Baldev, 1959; Митрофанова, 1961; Рудаков, 1961; Мае Leod, 1961; Терехова, 1962; Fujita, 1964, и др.).
Рассмотрим онтогенез Cuscuta еигораеа в качестве примера метаморфи- зованного способа развития в этой группе паразитных растений. Ко времени созревания семени зародыш приобретает морфогенетически униполярную организацию. Это означает, что способность к дальнейшему развитию сохраняет лишь один из полюсов зародыша — апикальный полюс. Зародыш в зрелом семени дифференцирован на эмбриодерму, основную ткань, более или менее сложно организованную васкулярную ткань и меристему стеблевого апекса. Подобный тип организации характерен и для других видов повилик (Johri, Tiagi, 1952; Fujita, 1964, и др.).
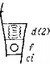
Базальный полюс зародыша Повиликовых представляет собой образование весьма оригинальное. Хацциус и Троль (Haccius, Troll, 1961) отмечали, что у зародышей повилик полностью отсутствует гипофиз. После отмирания суспензора на месте гипофиза остается открытое разрушенное место. Действительно, наши наблюдения показывают, что гистологическая организация банального полюса в зародышах повилик весьма своеобразна (рис. 20, 2, 5). Это своеобразие заключается в том, что базальная часть зародыша представляет собой продолжение эпикотиля. Какие- либо черты радикулярной организации здесь полностью отсутствуют, из чего можно заключить о полной редукции радикулы.
Прорастание повилики начинается с того, что базальный полюс зародыша прорывает размягченную оболочку и, обладая позитивной геотро- пичностью, проникает в верхний слой почвы, где и закрепляется. Вслед за этим из оболочки семени освобождается апикальный полюс зародыша; в это время проросток повилики начинает производить спиралеобразные ростовые движения.
Для нас важно отметить, что выход базального конца зародыша повилики из семени и его закрепление в почве основаны исключительно на ростовых явлениях, происходящих за счет растяжения клеточных оболочек и не сопровождающихся явлениями морфогенетического развития (рис.
После закрепления проростка на стебле растения-хозяина и развития первых присосок-гаусториев базальная область проростка постепенно отмирает. Таким образом, на первой стадии прорастания, на стадии поиска питающего субстрата, происходит униполярное развитие проростка (протосомы) повилики.
Возвращение проростка к биполярной организации в связи с переходом к половому воспроизведению происходит в результате вторичного заложения в теле протосомы гаусториальных и генеративных органов.
В противоположность представителям паразитных растений с первой формой паразитизма, униполярное развитие проростка повилики происходит за счет развития апикального (а не базального, как в случае с зара- зихами) полюса зародыша. В отличие от зародышей заразих, где полностью элиминируется эпикотиль, но сохраняются морфогенетические потенции ба-
зального (радикулярного) полюса зародыша, у повилик полностью элиминируется радикула, а морфогенетические потенции сохраняются в эпико- тилярных структурах. Эти особенности развития и соответственно разные формы редукции у заразих и повилик обусловливаются различиями в происхождении этих двух форм паразитизма. Щ
 Из сказанного можно сделать вывод, что способ послесеменного развития (форма метаморфоза) выявляет нам форму редукции зародышей.
Из сказанного можно сделать вывод, что способ послесеменного развития (форма метаморфоза) выявляет нам форму редукции зародышей.Состояние первичной А-протосомы представлено у повилик чрезвычайно отчетливо в связи с тем, что у повилик в течение всего онтогенеза спорофита осуществляется поиск питающего субстрата (новых растений- хозяев). 5-состояние протосомы совершенно не выражено. Вегетативное размножение осуществляется у повилик посредством ветвления первичной Л-протосомы, в связи с чем мы можем выделить здесь (^состояние протосомы. Таким образом, для Cuscutaceae характерно свое оригинальное направление в эволюции протосомы, характеризующееся становлением А- и С-состояний.
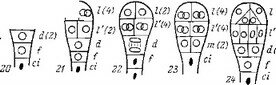
Рис. 22. Эмбриогенез некоторых представителей сем. Cuscutaceae.
japonica\ 13—15 — Cuscuta
еигораеа.
alt="" />
Еще по теме ПОСЛЕСЕМЕННОЕ РАЗВИТИЕ, МЕТАМОРФОЗ И ЭВОЛЮЦИЯ ПРОТОСОМЫ У CUSCUTA:
- ПОСЛЕСЕМЕННОЕ РАЗВИТИЕ И МЕТАМОРФОЗ У THUNIA MARSCHALLIANA
- ПОСЛЕСЕМЕННОЕ РАЗВИТИЕ И МЕТАМОРФОЗ У OROBAN С HE CREN AT А И CY NOMORIU M SONGARICUM
- ПОСЛЕСЕМЕННОЕ РАЗВИТИЕ И МЕТАМОРФОЗ У ПРЕДСТАВИТЕЛЕЙ СЕМ. PYROLACEAE
- ЭВОЛЮЦИЯ ПРОТОСОМЫ
- Часть 1. Развитие представлений об эволюции
- Глава 4 ЭВОЛЮЦИЯ И РАЗВИТИЕ
- 2-5. Развитие зародышей и эволюция. Жоффруа Сент-Илер
- Чайковский Ю.В. Наука о развитии жизни. Опыт теории эволюции., 2006
- МЕТАМОРФОЗ
- МЕТАМОРФОЗ, РЕДУКЦИЯ И НЕОТЕНИЯ И СИСТЕМЫ МОДУСОВ ЭВОЛЮЦИОННЫХ СТРУКТУРНЫХ ПРЕОБРАЗОВАНИЙ
- 14. Эволюция организмов и эволюция сообществ
- Эволюция популяций и эволюция организмов
- 6-1. Эволюция организмов и эволюция природы