ПОСЛЕСЕМЕННОЕ РАЗВИТИЕ И МЕТАМОРФОЗ У OROBAN С HE CREN AT А И CY NOMORIU M SONGARICUM
Одной из основных особенностей прорастания высокоспециализированных паразитных растений с первой формой паразитизма является униполярное развитие проростка из базального полюса зародыша (проэмбрио).
Изучение дальнейших этапов онтогенеза высокоспециализированных паразитных покрытосеменных растений привело нас к заключению, что наиболее общей и характерной их особенностью является развитие с метаморфозом организации спорофита. Метаморфоз, т. е. более или менее кардинальное преобразование организации индивида в процессе его* онтогенетического развития, известен до настоящего времени только* у представителей животного царства. Известно также, что метаморфоз: у представителей животного мира характеризуется прежде всего «превращением одной формы в другую, резко отличную» (Шмидт, 1951). Подобного рода превращения выражаются в изменении «внешнего вида и строения животного в послезародышевом развитии до достижения ими половозрелого состояния» (Павловский, 1960).
Морфогенетические преобразования, обозначенные Е. Н. Павловским как изменения в строении живых существ, означают по существу изменения таких основных аспектов структурной организации индивида, как полярность, симметрия, изменение характера, числа и соотношения частей и органов.
Эти изменения обусловлены изменениями в направлении и активности морфогенетических процессов. К этим преобразованиям относятся и смена очагов развития (исчерпывание старых и заложение новых, вторичных меристематических зон, имагинальных дисков), и некроз соматических зон, исчерпавших свои морфогенетические потенции, с использованием освобождающихся биопродуктов для дальнейшего развития индивида. Важно отметить, что речь идет при этом не о преобразовании отдельных частей или органов индивида, но о принципиальной перестройке его организации в целом. Фактически при этом происходит полное замещение одного типа организации индивида другим.Метаморфоз паразитных покрытосеменных растений, как и метаморфоз у животных организмов, также заключается в замене одного типа организации другим, принципиально отличным. Весьма существенны, на наш взгляд, для понимания явлений метаморфоза представления Берлезе
(Berlese, 1913; цит. по: Ежиков, 1929), развитые далее у нас И. И. Ежиковым (1929, 1936, 1939), а в Англии Иммсом (Imms, 1936). Согласно И. И. Ежикову (1929), основным и решающим фактором происхождения Holometa- bola необходимо считать более раннее вылупление зародыша из яйца, в результате которого личинка сохраняет в основном эмбриональную организацию (личинка есть свободноживущий зародыш, претерпевший обширные ценогенетические изменения).
Некоторые аспекты этих представлений были подвергнуты критике (Гиляров, 1949, 1957; Hinton, 1953, 1955), однако основное ядро теории (идея о гомологии зародыша и личинки) сохраняет свое значение и сегодня. Наши материалы подтверждают идею Берлезе—Ежикова—Иммса на примере паразитных цветковых растений. Конечно, конкретное морфогенетическое выражение явлений метаморфоза и признаков зародыша у «свободноживущих протосом» («личинок») спорофита паразитных растений может быть сколь угодно отличным от подобных явлений в мире Holometabola, однако в общебиологическом плане сходство способов эволюционных преобразований онтогенеза у животных и у паразитных цветковых совершенно очевидно.
В онтогенезе высокоспециализированных паразитных цветковых растений метаморфоз находит свое выражение прежде всего в переходе от униполярного к биполярному способу развития спорофита. Это явление происходит, естественно, на основе изменений в фундаментальных структурах, обеспечивающих то или иное течение морфогенетических процессов (Терёхин, Никитичева, 1968; Терёхин, 19686).
Метаморфоз в онтогенезе животных всегда обусловлен более или менее резкими изменениями образа жизни в течение индивидуального развития. Часто это связано со сменой среды обитания, например с переходом от планктонного образа жизни к сидячему у губок, мшанок, кишечнополостных и асцидий, с переходом от наземного или водного образа жизни к воздушному у многих насекомых, с выходом из воды на сушу у амфибий, с переходом к паразитному образу жизни у многих червей, ракообразных, некоторых клещей и т. д. (Lameere, 1900; Мартынов, 1938; Snodgrass, 1944; Захваткин, 1949; Шванвич, 1949; Шмидт, 1951; Chen, 1955; Гиляров, 1957; Расницын, 1965; Токин, 1966, и др.). Изменения в образе жизни, связаны ли они с переходом к паразитизму или со сменой среды обитания, означают прежде всего изменения в функциональной активности организмов (Гиляров, 1957).
Общую картину функциональных и морфогенетических изменений, обусловленных переходом от автотрофного к паразитному существованию у покрытосеменных растений можно представить себе следующим образом. Как известно, по выходе из семени проросток автотрофного растения (например, Verbascum) переходит к самостоятельному, независимому от материнского растения, существованию, и в этот период его основная задача состоит в осуществлении вегетации, возможность которой обеспечивается, помимо прочего, и соответствующей структурной организацией. Вегетация у автотрофного растения реализуется посредством двух основных систем: корневой и стеблелистовой.
Иная картина предстает перед нами при анализе жизнедеятельности проростка высокоспециализированного паразитного растения.
Вместо двух источников питания из почвы и воздушной среды проросток паразитного растения, перешедший к чужеядному питанию, имеет единственный источник пищевых материалов — растение-хозяина. Очевидно, что при этой ситуации биполярная организация растения приходит в противоречие с новым образом жизни. Вследствие дискретного распределения питающего субстрата (растений-хозяев) растение-паразит, выйдя из семени, осуществляет функции поиска и освоения питающего субстрата не сопряженно, что характерно для автотрофных растений, Ho последовательно: сначала идет поиск питающего субстрата, а затем его освоение. И то и другое реализуется посредством одной структуры, поскольку источник питания один — растение-хозяин. Несомненно, что при этой ситуации биполярность организации биологически неоправданна.
Развитие зародыша-проростка специализированного паразитного растения в период поиска и освоения питающего субстрата (растения-хозяина) происходит униполярно. Поскольку, однако, для любого паразитното растения остается незыблемой необходимость полового воспроизведения и поскольку последнее осуществляется теми же функциональными и структурными средствами, что и половой процесс у автотрофных покрытосеменных растений, у растений-паразитов при наступлении половой репродукции должен происходить метаморфоз организации, возвращение ее к биполярной системе: органы половой репродукции—вегетирующая питающая система.
Таковы фундаментальные изменения, происходящие в функциональной и структурной организациях покрытосеменных растений в процессе эволюции паразитизма. В явлении метаморгфоза, очевидно, мы находим как бы компромисс между необходимостью все более экономичного осуществления нового способа питания (в тканях растения-хозяина, униполярно) и необходимостью сохранения старого способа полового воспроизведения в воздушной среде (отсюда вторичное для онтогенеза становление биполярной организации).
Среди зоологов, изучающих эволюцию животных с метаморфозом, идея структурного преобразования онтогенеза под воздействием дивергенции в эволюции функций (образа жизни), обусловленной существованием в различных средах, находит вполне определенную поддержку (Snodgrass, 1944; Гиляров, 1957, и др.).
Каким же образом происходит послесеменное развитие высокоспециализированных паразитных растений в группе с первым способом паразитизма?
Интерес к изучению онтогенеза у паразитных растений из группы ксенопаразитов с первым способом чужеядности пробудился, по-видимому, в начале второй половины прошлого века, после того, как Шахт (Schacht, 1850) впервые правильно описал слабодифференцированные зародыши в зрелых семенах заразих. Он описал эти зародыши как овальные, сравнительно немногоклеточные образования без дифференциации на семядоли, радикулу, эпикотиль и гипокотиль. Тогда же Каспари (Caspary, 1854) впервые описал развитие проростка заразихи (Orobanche ramosa) на корнях конопли. Каспари отметил, что вначале у зародыша развивается базальный (корневой) полюс; несколько позже проросток значительно утолщается в месте внедрения в ткани корня растения-хозяина. На образовавшемся «клубеньке» прежде всего возникают дополнительные «корешки», и лишь вслед за этим в апикальной области клубенька появляются две первые чешуи, в которых Каснари признал запаздывающие в своем развитии семядоли.
Слабо дифференцированные зародыши и характерные бульбообразные проростки были вскоре обнаружены у других заразих (например, Epi- fagus), а также у паразитов из семейств Balanophoraeeae и Cynomoriaeeae (Hooker, 1855; Weddel, 1858—1861; Eichler, 1869).
Веддел (Weddel, 1858—1861) отчетливо показал, что у Cynomorium coccineum при прорастании развивается лишь базальный полюс слабодифференцированного зародыша, в то время как его апикальная область не обнаруживает сколько-нибудь заметных морфогенетических изменений. Дальнейшие исследования высокоспециализированных паразитных растений (в основном изучались представители сем. Orobanchaceae в связи с их практической значимостью) подтвердили униполярный характер развития молодых проростков этих растений (Koch, 1877b, 1887; Kusano, 1908; Heinricher, 1910; Juliano, 1935; Rauh, 1937; Бейлин, 1947а, 1968; Kadry, Tewfic, 1956а, 1956b).
Неясным оставалось происхождение и судьба стеблевого апекса (эпикотиля).
Используя оригинальную методику, египетские исследователи Кадри и Тьюфик (Kadry, Tewfic, 1956а, 1956Ь) изучали проростки заразихи Orobanche crenata. Они пришли к выводу, что из базального-(корневого) полюса зародыша заразихи в процессе прорастания развивается «трубкообразный орган» («germ tubelike organ»), по строению принципиально отличный от радикулы. Этот трубкообразный проросток проникает в ткани корня растения-хозяина и там формирует гаусториальный орган, а снаружи корня формируется «клубенек». В клубеньке эндогенно образуются апекс побега и многочисленные корнеподобные органы. К сожалению, эти выводы не были подкреплены анатомическими картинами, действительное морфогенетическое происхождение стеблевого апекса оставалось неясным.В 1958 г. Креннер (Krenner, 1958) на примере Orobanche подтвердил представление Коха (Koch, 1878) о том, что эпикотиль дифференцируется из клеточных слоев зародыша, противоположных по своему положению тем, из которых развивается гаусторий.
В связи с существующими разногласиями мы избрали для изучения по- слесеменного развития высокоспециализированного паразитного растения с первой формой паразитизма тот же вид заразихи, который ранее был изучен Кадри и Тьюфиком, — Orobanehe crenata. Использовав для получения проростков методику этих авторов, мы изучали (Терёхин, 19686; Терёхин, Никитичева, 1968) поэтапно не только морфологию, но и анатомическое строение проростков (табл. III; рис. 9, 10)»
Зародыш в зрелом семени О. crenata представлен немногоклеточным глобулярным слабодифференцированным телом, в котором можно ясно различить лишь клетки эмбриодермы и основной меристемы (рис. 9,1). В зародыше можно также различить слабую полярную дифференциацию: клетки апикальной области зародыша несколько крупнее, чем базальной, размеры клеток уменьшаются постепенно.
С началом прорастания клетки базальной области проэмбрио О. crenata активируются под влиянием выделений из корней растения-хозяина и быстро пролиферируют (в основном за счет процессов направленного клеточного роста), образуя вскоре, по терминологии Кадри и Тьюфика, «трубкообразный проросток», в котором ни с какой стороны нельзя признать образование, анатомически или морфологически подобное первичному корню (радикуле). «Трубкообразный проросток, — отмечают Кадри и Тьюфик (Kadry, Tewfie, 1956а, стр. 279), — не может быть признан за радикулу, так как имеются большие различия в их анатомических и морфологических структурах. Он скорее подобен прорастающей трубке грибной споры». Действительно, анатомия проростка в этот период крайне проста. Он состоит из однородных тонкостенных, паренхиматозных клеток, без дифференциации на эпидермис или васкулярную ткань (рис. 9, 2, 3).
Трубкообразный проросток растет в направлении корня растения- хозяина. После контакта с корнем растения-хозяина клетки терминальной области проростка активно внедряются в ткани корня в направлении центрального цилиндра, в то время как ближайшая к корню часть проростка диффузно разрастается, образуя в конце концов «клубенек» (рис. 9, 10, I—4). Апикальная область проэмбрио, остающаяся в семени, далее не развивается, но ее клетки, выполнив гаусториальную функцию в эндосперме, в дальнейшем отмирают. Клетки апикальной области проэмбрио, выполняющие гаусториальную функцию, отчетливо различимы уже с самых первых этапов прорастания (рис. 9, 2). Вслед за апикальной областью проэмбрио, после внедрения проростка паразита в ткани корня хозяина и образования клубенька, отмирает и средняя часть сомы проростка. Содержимое отмирающих клеток используется для построения клубенька- протосомы (рис. 10, 3). 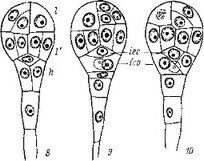 Э. С. Терёхин
Э. С. Терёхин
Рис. 9. Послесеменное развитие Orobanche crenata (сем. Orobanchaceae).
Первая фаза; I *¦ зрелое семя; 2, 3 — начало прорастания; 4 — внедрение в ткани растения*
хозяина.
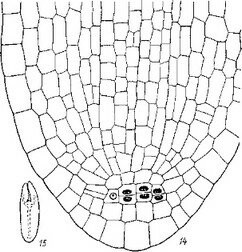
Рис. 9 (продолжение).
Из приведенных выше описаний, рисунков и фотографий достаточно ясно, что первоначальное развитие проростка (развитие внесеменной стадии зародыша, стадии протосомы) происходит униполярно: развивается только базальная область зародыша, в то время как его апикальная область утрачивает постепенно, начиная со времени прорастания, свое функциональное значение и отмирает.
Образование гаустория и клубенька — это морфогенетическое выражение процесса освоения пищевого субстрата и одновременно подготовка к выполнению очередной функциональной задачи — развитию воспроизводящей половой системы. На сравнительно ранней стадии развития клубенька («шишечки», «бульбы», «каллуса», по терминологии некоторых авторов) в его терминальной' области эндогенно, под отмирающими остатками семени, можно наблюдать процесс возникновения сравнительно многоклеточного меристематического очага, формирующегося из однородных паренхиматозных клеток, составляющих тело протосомы (рис. 10, i, 2). Этот очагзахва-
©\lt;sgt;
Рис. 10. Послесеменное развитие Orobanche crenata (сем. Orobanchaceae). Вторая фаза: I—5 — эндогенное образование апекса генеративного побега.
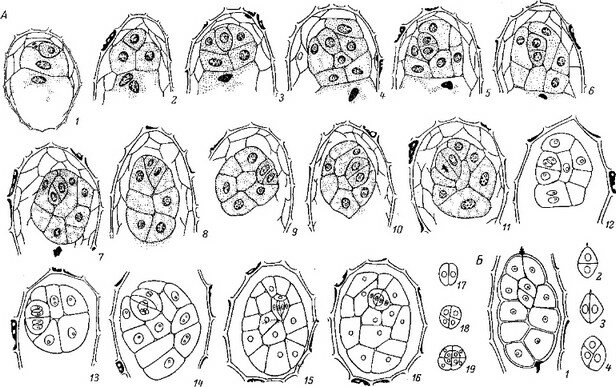
Рис. 10 (продолжение).
тывает вначале*примерно 3—8 слоев клеток. Характернейшая черта этого процесса состоит в том, что эпидермальный слой клеток вновь возникшего меристематического очага вскоре резко отграничивается от остающихся кнаружи от него паренхиматозных клеток, содержимое которых быстро расходуется на построение структур вновь возникшего очага меристема- тической ткани (рис. 10, 1, 2). Напротив, внутренняя граница меристематического очага остается нечеткой: меристематические клетки постепенно переходят в паренхиматозные (рис. 10, 2—4).
Довольно скоро происходит и пространственное отделение эпидермального слоя клеток меристематического очага от лежащих кнаружи паренхиматозных клеток. Образуется щель, по краям которой активно продолжается процесс меристематизации (рис. 10, 4). Отдельные области, расположенные латерально от основного меристематического очага, разрастаются более активно; клетки в них, как мы уже отметили, становятся все более меристематическими. Проследив дальнейшее развитие меристематического очага, можно видеть, что если его центральная область развивается в дальнейшем в стеблевой апекс, то вышеупомянутые латеральные его выросты суть первые чешуи, прикрывающие этот вновь образующийся апекс генеративного побега (рис. 10, 5). Таким образом, мы устанавливаем, что происхождение первых и следующих чешуй морфогенети- тически не гомологично. Возникает вопрос, можно ли назвать эти первые кроющие чешуи семядолями? Известно, что семядоли автотрофных
растений возникают в одно время со стеблевым апексом или несколько раньше из латеральных областей проэмбрио, в то время как настоящие листья — производные стеблевого апекса. В отличие от семядолей автотрофных растений чешуи, возникающие в протосоме О. crenata не выполняют ни гаусториальной, ни фотосинтезирующей, ни, вероятно, запасающей функции. Основная функция чешуй — предохранение стеблевого апекса от повреждений при прохождении побега через ночву.
После сформирования стеблевого апекса клубенек представляет собой биполярное (побег—гаусторий) образование. Генеративных побегов, равно, как и вторичных гаусториальных систем, в одной бульбе может возникнуть несколько. Как показал Pay (Rauh, 1937), зачатки генеративных побегов могут появиться в разных частях протосомы, но первый из них (на это ясно указывают наблюдавшиеся нами картины) дифференцируется, как правило, непосредственно под отмирающими частями трубкообразного проростка, а остальные — в прилегающих апикально-латеральных областях.
Почти одновременно с образованием зачатков генеративного побега, а иногда и несколько раньше в теле протосомы, в латеральных ее областях, возникают многочисленные вторичные протосомы, которые по своей внутренней структуре более подобны корням, нежели первичцая протосома, но существенно от них отличаются отсутствием чехлика и корневых волосков. Некоторые исследователи как будто находили образования, подобные корневому чехлику, на этих вторичных протосомах заразих (Koch, 1887; Smith, 1904). Однако ряд других исследователей, особенно исследователи новейшего времени, говорят об отсутствии каких-либо структур, подобных чехлику (Worsdell, 1895; Ghemin, 1920; Juliano, 1935; Kadry, Tewfic, 1956а). В пользу отсутствия чехлика свидетельствуют и наши наблюдения.
Основная функция вторичных протосом — вегетативное размножение. Приходя в контакт с корнями растения-хозяина, они образуют вторичные присоски-гаустории; питательные вещества, производимые этими последними, идут вначале, по-видимому, на питание основной протосомы, а затем на формирование вторичных каллусов. Весь процесс развития системы гаусторий—побег повторяется снова. Образование васкулярных тканей начинается в клубеньке довольно рано, одновременно с началом образования вторичных меристематических очагов. Васкулярная система клубенька уже в самом начале своего развития связывает меристематические центры с активно работающими клетками присоски (гаустория).
Подведем краткие итоги. Внутрисеменной период развития зародыша резко сокращен, он заканчивается на стадии позднего (глобулярного) проэмбрио. В первой послесеменной период развития (стадия «трубкообразного проростка» протосомы) проросток-протосома развивается униполярно из клеток базальной области проэмбрио, что наилучшим образом отвечает задаче поиска питающего субстрата (растения-хозяина), единственного источника пищевых ресурсов растения-паразита. Во второй послесеменной период развития (стадия «клубенька») происходит диффузное сферическое разрастание тела проростка-протосомы и затем возвращение к биполярной системе развития за счет возникновения вторичных меристематических очагов («имагинальных дисков»), из которых развиваются генеративные побеги.
Смена униполярного биполярным способом развития характерна и для других высокоспециализированных паразитных цветковых (рис. 11)
Сравнив развитие какого-либо автотрофного растения (например, Veronica) и развитие Orobanche1 нельзя не прийти к выводу, что характер морфогенетических процессов в онтогенезе Veronica столь же хорошо соответствует ее автотрофному образу жизни, сколь точно соответствует метаморфизованный онтогенез заразихи образу жизни высокоспециализированного паразита.
Описанные выше процессы морфогенеза в проростке-протосоме мы наблюдали и у другого представителя высокоспециализированных ксено-
Рис. И. Онтогенетическое развитие с метаморфозом у некоторых высокоспециализированных паразитных цветковых растений (схематизировано).
А — автотрофныйЦVerbascum thapsus: I—6 — внутрисемейное развитие зародыша; 7 — проросток. Б — Orobanche crenata: I—3 — внутрисемейное развитие зародыша; 4, 5 — внесеменное униполярное развитие зародыша; 6,7 — переход|к биполярному развитию; 8 — проросток. В — Hypopi- tys monotropa: I — зрелое семя; 2—4 — внесеменное униполярное развитие зародыша; 5,6 — проростки. Г — Thunia marschalliana: I—4 — внутрисемейное развитие зародыша; St 6 — внесеменное униполярное развитие зародыша; 7 — проросток. Д — Cuscuta еигораеа: I—4 — внутрисемейное развитие^зародыша;1гgt;, 6 — внесеменное униполярное развитие зародыша; 7 — проросток.
паразитов — у Cynomorium songaricum. Веддел (Weddel, 1858—1861) еще в прошлом веке показал, что зародыш-проросток С. coccineum развивается в период поиска растения-хозяина униполярно из базальной области зародыша. Изученные нами картины развития протосомы у С. songaricum показывают их принципиальное тождество с развитием протосомы у Orobanche crenata. Для С. songaricum также характерно заложение генеративного побега за счет образования эндогенных меристематических очагов (табл. IV, рис. 12).?
ЭВОЛЮЦИЯ ПРОТОСОМЫ
Пути развития протосомы у паразитных цветковых с первой формой паразитизма могут быть довольно различны. Однако они имеют и одну общую черту: протосома всегда развивается из базального (корневого)

Рис. 12. Послесеменное развитие Cynomorium songaricum (сем. Cynomoriaceae).
I—6 — последовательные этапы эндогенного образования апекса генеративного побега, ап — апекс генеративного побега; прч — кроющие чешуи; к — корень растения-хозяина.
полюса зародыша. Это обстоятельство имеет важное значение, так как приводит к выводу о единой форме редукции зародышей у растений с одним способом чужеядности, с одной формой паразитизма. Сохранение морфогенетической активности базального полюса зародыша при полном угнетении такого рода активности на апикальном его полюсе — таковы основные черты этой формы редукции.
Рассмотрим некоторые вопросы эволюции протосом на примере конкретных паразитных семейств.
Становление протосомы у представителей сем. Норичниковых совпадает со временем завершения редукции фотосинтезирующего аппарата (рис. 13, В). У высокоспециализированных Норичниковых (Harveya, Hyobanehe9 Striga orobanehoides) представлены все основные (A9 B9 С) состояния протосомы (табл. 2). При этом С-протосома представлена в виде С2-состояния (т. е. вегетативное размножение протосомы происходит здесь посредством образования вторичных .4-состояний). Для понимания эволюции протосомы в сем. Норичниковых наиболее перспективно изучение онтогенеза у представителей рода Striga. Проростки Striga euphrasioides и S. elegans (Marloth, 1934; Tiagi, 1956) слабоспециализированные паразиты с биполярным
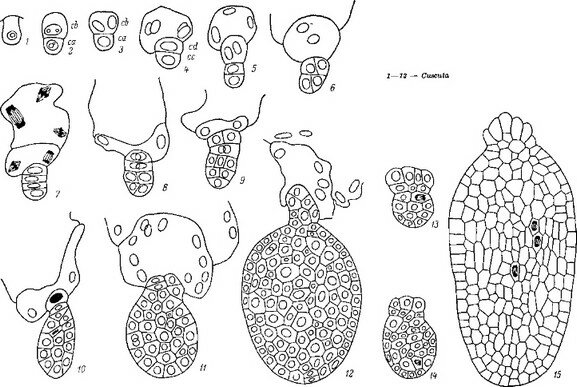
Рис. 13. Послесеменное развитие некоторых паразитных представителей сем. Scro- phulariaceae с разной степенью морфогенетической адаптации к паразитному образу
Pжизни (схематизировано).
А — Euphrasia; В — Lathraea; В — Striga orobanehoides', Г — Hyobanche. I — зародыш в зрелом семени; 2,3 — проростки; 4 — цветущий побег.
/>способом развития. С другой стороны, у S. orobanchoides мы обнаружили вполне сложившееся типичное метаморфизованное развитие, сходное в своих основных чертах с метаморфозом Orobanche crenata.
Некоторые виды Striga дают примеры переходных форм. Марло (Mar- loth, 1932) отмечал, что добавочные корни из подземной части побега
S. Iutea участвуют в паразитизме, образуя присоски на корнях растений- хозяев. Мы обнаружили во множестве такие же корнеподобные образования на подземных частях старого побега S. orobanchoides. В то же время сам каллус (клубенек, 5-состояние протосомы) у S.} orobanchoides не несет на себе органов вегетативного размножения. Однако небольшого смещения времени заложения адвентивных корневых структур вполне достаточно, чтобы мы увидели перед собой типичную картину С2-состояния протосомы характерную для многих Заразиховых и, вероятно, для Harveya и Hyobanche,
Несомненно, что возникновение клубенька как места накопления и хранения веществ необходимых для формирования генеративного побега является образованием de novo, не имеющим прецедента в предшествующей истории эволюции автотрофных Норичниковых. Идентификация стеблевого апекса, возникающего в клубеньках эндогенно, не представляет затруднений. Здесь нетрудно распознать адвентивную природу побега, закладывающегося в тканях клубенька. Таким образом, в процессе эволюции протосомы происходит de novo формирование клубенька и замещение (субституция) основного генеративного побега (развивающегося у автотрофных растений из эпикотиля) адвентивным, формирующимся вторично и эндогенно в тканях клубенька.
Процесс дифференциации генеративного побега, подобный тому, что был описан выше на примере Orobanche crenata, характерен, по ^ашим наблюдениям, и для Striga orobanchoides.
Мы уже отмечали несогласие исследователей в вопросе о судьбе апикального полюса зародыша у заразих. Так, Kox (Koch, 1887) вслед за Каспари (Caspary, 1854) утверждал, что семядоли и стебель проростка заразих развиваются из апикального полюса зародыша. Pay (Rauch, 1937) считала что у Orobanche speciosa наблюдается обычное для семядольных растений развитие стеблевого полюса зародыша, тогда как у О. hederae апикальная область зародыша всегда отмирает и побеги развиваются адвентивно из тела клубенька (каллуса). Кадри и Тьюфик (Kadry, Tewfic, 1956а, 1956Ь) дали в общем правильное, но, к сожалению, очень неполное морфологическое описание первых этапов прорастания заразих. Креннер (Krenner, 1958) присоединился к точке зрения Коха, а Ранган и Рангасвами (Rangan, Rangaswamy, 1968) подтвердили данные Кадри и Тьюфика.
Детальное анатомическое исследование послесеменного развития заразих, проведенное нами на примере О. crenata (Терёхин, Никитичева, 1968) позволило дать более или менее полное представление об этой форме метаморфоза. То, что нам известно сегодня об уровне и особенностях дифференциации зародышей в зрелых семенах других Заразиховых и сведений (хотя и очень фрагментарных) об их послесеменном развитии, достаточно, по-видимому, для утверждения, что метаморфоз характерен для всех Заразиховых.
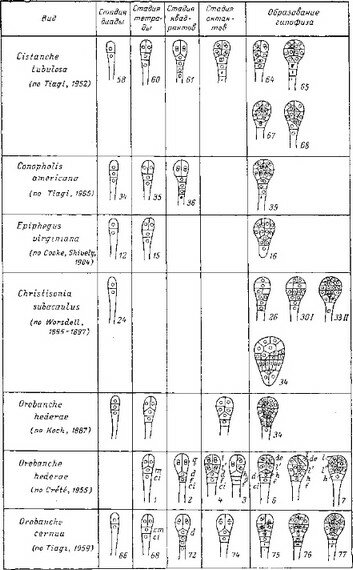
Очевидно, для всех заразих характерно отчетливое Л-состояние протосомы, развивающейся из базального полюса зародыша. Существование 5-протосомы подтверждено для всех исследованных к настоящему времени представителей семейства (Caspary, 1854; Koch, 1887; Wilson, 1904; Boeshore, 1920; Rauh, 1937; Бейлин, 1950а; Kadry, Tewfic, 1956а, 1$56Ь; Терёхин, Никитичева, 1968, и др.).
Изменение С-состояния протосомы в пределах сем. Заразиховых весьма значительно. С-состояние протосомы является структурным выражением способа вегетативного размножения. В пределах семейства можно от
четливо различить два пути осуществления этой задачи. В первом случае задача решается «почкованием» .В-протосомы (т. е. заложением дополнительных меристематических очагов развития генеративных побегов). Эта тенденция намечается у некоторых видов Orobanche (О. epithymum) (табл. 2) и весьма выражена в родах Conopholis, Epiphagus и Cistanehe (Wilson, 1904; Boeshore, 1920; Бейлин, 1950, и др.). Мы обозначаем такой способ развития как ^-состояние протосомы.
Во втором случае задача вегетативного размножения осуществляется за счет образования вторичных Л-протосом, т. е. преобразованных в гаустории корней, которые, выйдя из материнской 2?-протосомы, прикрепляются к телу хозяина и образуют новые, вторичные Б-протосомы (Rauh, 1937; Kadry, Tewfic, 1956Ь; Терёхин, Никитичева, 1968). Этот способ развития, обозначаемый нами как С2-состояние протосомы, характерен для Orobanehe erenata, О. hederae, О. ramosa, О. flava, Phelypaea coeeinea, Aeginetia indica (табл. 2). Модификацией и дальнейшим усовершенствованием этого способа развития может служить развитие интраматрикаль- ной системы гаусториев у Orobanche minor, О. rapum, О. hederae (Rauh, 1937).
У Phacellanthus tubif Iorus, с другой стороны, нами обнаружено типичное ^-состояние протосомы, при котором задача вегетативного размножения реализуется за счет ветвления А-протосомы. В некоторых случаях (О. epi- thimum и др.) сопряженно развиваются две системы вегетативного размножения (Rauh, 1937).
Таким образом, для эволюции протосом Заразиховых характерны три самостоятельных направления.
Для всех представителей сем. Баланофоровых также характерно развитие зародышей-проростков (протосом) из базального полюса проэмбрио. А- и 5-состояния протосомы хорошо выражены. В эволюции способов вегетативного размножения у членов семейства можно выделить две основные тенденции: развитие системы почкования (^-состояние протосомы) и системы ветвления Л-протосом (С3-состояние протосомы). Наиболее простые формы вегетативного размножения мы находим у представителей родов Scybalium, Lathrophytum, Lophophytum, Ambrophytum, Sarcophyte и Mistropetalon. У этих растений почкование Б-протосомы выражено слабо.
Среди видов Balanophora мы находим такую, например, форму, как Balanophora ungeriana, которую Pay (Rauh, 1937) рассматривает в качестве переходной от Orobanche hederae к более специализированным видам Balanophora (В. globosa). Если у В. ungeriana дополнительные генеративные побеги возникают путем почкования только после развития главного, терминального (но адвентивного в сущности!) побега, то у В. glodosa все генеративные побеги образуются почти одновременно. 5-протосомы обоих вышеназванных видов обнаруживают сильное развитие системы почкования. В. elongata не представляет собой, по-видимому, переходной формы к системам вегетативного развития Langsdorffia и Thonningia, как думает Pay (Rauh, 1937), мнение которого раньше разделяли и мы (Терёхин, Никитичева, 1968). По-видимому, правильнее говорить о параллельной, но относительно независимой эволюции систем вегетативного размножения этих растений в границах семейства.
Изучение послесеменного развития Cynomorium songaricum (Cyno- moriaceae) показывает, что у этого паразита хорошо выражены все три (А, В, С) состояния протосомы. При этом вегетативное размножение происходит, как и у * Заразиховых, за счет образования вторичных А-протосом (С2-состояние протосомы). Можно заключить отсюда, что эволюция послесеменного этапа развития спорофита Cynomorium происходила в том же направлении и теми же способами, что и эволюция протосомы в сем. Orobanchaceae.
Строение зародышей в зрелых семенах Pholisma и Lennoa (Lennoaceae)9 строение самого семени и данные о характере развития спорофита после перехода к чужеядности (Britton, 1930; Schmucker, 1959b; Kuijt, 1966, 1967; наши наблюдения) свидетельствуют о том, что и здесь развитие паразита происходит из базальной области проэмбрио. Хорошо выражены A-, В- и С2-состояния протосом. С^-состояние протосом усложнено у Len- поасеае и Hydnoraceae явлением диморфизма вторичных протосом (Kuijt, 1969).
^-состояние протосомы у представителей сем. Hydnoraeeae выражено очень слабо. Вегетативное размножение первичной протосомы основано, по нашим наблюдениям, на ветвлении Л-протосомы. По способу вегетативного размножения протосомы представителей сем. Hydnoraeeae близки к Phaeellanthus tubiflorus из Orobanehaeeae. Для проростков обоих растений характерны также частные анастомозы между отдельными ветвями протосомы.
Изучение послесеменного развития у представителей сем. Ремнецветниковых показало, что в семействе представлены как переходные формы от прямого автотрофного Способа развития спорофита к метаморфизо- ванному развитию специализированного паразитного растения, так и формы с метаморфизованным развитием. Сравнительно более полно по- слесеменное развитие в этой группе было изучено у Viseum album (Tubeuf, 1923; Rauh, 1937) и Loranthus eelastroides (Me Luckie, 1923). Вот как описывает прорастание Viseum album Тубеф (Tubeuf, 1923, стр. 465—466): «В ягоде Viseum лежит I—4 зародыша. Каждый из них развивается самостоятельно. При прорастании гипокотиль выходит из остатка семени, изгибается к поверхности ветви растения-хозяина, к которой прикреплено так называемое семя (в действительности плод, — Э. Т.) и образует на корневом полюсе зародыша липкую „шайбу“, которая крепко прикрепляется к подстилающей ветви. В это время у зародыша оба полюса фиксированы: вершина побега остается в прилипшей ягоде, а другой конец образован липкой „шайбой". В этом состоянии зародыш обычно перезимовывает. Следующей весной ему удается своим первичным гаусторием, который развивается из центральной части липкой шайбы, придти в контакт с проводящим пучком в тканях растения-хозяина. Тогда зародыш начинает выпрямляться. Он вытаскивает свой верхний конец из ягоды, или же поднимает последнюю вверх, поскольку она к тому времени теряет липкость. На освободившемся таким образом зародыше-проростке можно различить оба остатка семядолей и пару маленьких, лежащих наперекрест семядолям, первых листочков, расположенных непосредственно над семядолями или между ними. Летом эти первые листочки развертываются, иногда же перезимовывают в еще сложенном положении». На следующий год проросток развивает первые побеги из почек или обычно из одной почки, расположенной в пазухе первого листа.
Согласно Pay (Rauh, 1937), у юного растения Viseum album развивается в первый год только гипокотиль. На втором году закладываются стеблевые почки. Семядоли служат сосущими органами и к этому времени отмирают. Развитие стебля состоит в том, что первые почки, стоящие ближе всего к семядолям, образуют первые листья. На третий год развиваются побеги.
Mak-Люкки (Me Luckie, 1923) также отметил отмирание семядолей и эпикотиля и развитие побегов из адвентивных почек у Loranthus eelastroides.
Таким образом, у Viseum album и Loranthus eelastroides мы обнаруживаем переходное состояние к развитию с метаморфозом. Хотя здесь и присутствуют развитые семядоли и эпикотиль, последний не функционирует обычным образом. Растения развиваются из адвентивных меристем. Базой для развития адвентивных структур являются ткани эпикотиля. Это еще не метаморфоз, но уже и не прямое биполярное развитие.
Следующая ступень эволюции послесеменного развития спорофита
Ремнецветниковых может быть прослежена на примере Arceuthobium campulopodum. Как отметил Коэн (Cohen, 1963, 1965), стеблевой апекс зародыша этого растения вполне рудиментарен и в процессе прорастания, после внедрения проростка в ткани растения-хозяина, отмирает вместе с остатками всей апикальной области проростка. Здесь перед нами пример типичного метаморфизованного способа развития. У Phrygilanthus aphyllus Рейхе (Reiche, 1904) также обнаружил отмирание всех экстра- матрикальных частей проростка после внедрения паразита в ткани хозяина. Как видно, и здесь перед нами типичный случай метаморфизованного развития.
Говоря об эволюции различных состояний протосомы, мы должны отметить постоянное присутствие и прекрасную выраженность Л-состояния протосомы, характерного для всех изученных представителей семейства. 5-состояние протосомы вполне выражено только у немногих представителей семейства, не обладающих развитым аппаратом вегетативного размножения (табл. 2), например у Viscum articulatum. У видов с более или менее развитыми экстраматрикальными (Loranthus micranthus) или ин- траматрикальными (Phrygilanthus aphyllus) аппаратами вегетативного размножения 5-состояние протосомы выражено слабо или вообще не выражено.
Для представителей сем. Ремнецветниковых характерно наличие двух систем вегетативного размножения протосомы: экстраматрикальной системы «воздушных корней» и интраматрикальной системы «коровых корней» («коровых лучей», «эндофитной системы»).
В сем. Ремнецветниковых можно наблюдать, по всей вероятности, лишь одну основную тенденцию в эволюции С-протосом — тенденцию к смене экстраматрикальной системы вегетативного размножения (устроенной по типу создания вторичных Л-протосом) системой интраматрикальной (развитие эндофитной системы). К растениям, обладающим экстраматрикальной системой вегетативного размножения, из Ремнецветниковых можно, вероятно, отнести уже упомянутый выше Loranthus micranthus, а также в основном тропические растения Oryctanthus ruficaulis, О. occidentalism Phtirusa pyrifolia, некоторые виды Struthantusi Notothixos Incanusi Lepidoceras aquamifer, Eubrahion ambiguum и т. д. (Menzies, 1954; Thoday, 1956, и др.).
От переходных форм Viscum articulatum и V. capense через V. Orientalei У. cruciatum и далее через V. rotundifolium со слабо развитой эндофитной системой, не производящей генеративных побегов, мы можем перейти к целой группе видов, у которых наряду с формированием адвентивного экстраматрикального побега из первичного клубенька-гаустория хорошо развит и аппарат интраматрикального распространения паразита и интраматрикальное же производство адввнтивных генеративных побегов. Сюда относятся уже известный нам Viscum album, а также V. Crassulaei некоторые виды Loranthus и, вероятно, также Arceuthobium Occidentalei у которого первичные адвентивные побеги происходят из тела клубенька н далее Korthalsella salicornioides и К. opuntia.
Наряду с Phrygilanthus aphullus и Arceuthobium minimum, у которых экстраматрикально генеративные побеги вообще не образуются, а 5-состояние протосомы еще более угнетено, мы встречаем у некоторых других представителей последнего рода (А. minutissimum, А. americanum) и систему изофазного развития паразита и хозяина (Reiche, 1904; Thoday, 1956; Schmucker, 1959b; Kuijt, 1960; Cohen, 1965, и др.). У других видов Arceuthobium (A. douglasiii А. campylopodum), а также у представителей других родов сем. Loranthaceae изофазного развития обнаружено не было (Kuijt, 1960). Способностью к изофазному развитию обладают только отдельные виды сем. Ремнецветниковых, принадлежащие к группе растений с наиболее развитой системой интраматрикального вегетативного размножения, т. е. наиболее специализированные растения.
Таким образом, общее направление эволюции послесеменного развития у Ремнецветниковых можно обозначить как тенденцию к усилению интра- матрикальности и изофазности их развития.
Послесеменное развитие Раффлезиевых изучено еще недостаточна полно. Для всех исследованных видов характерен метаморфизированный способ послесеменного развития спорофита. По-видимому, для всех Раффлезиевых характерна инвазия в ткани растения-хозяина в почвенной или пограничной (на поверхности почвы) среде. Отсюда очевидно, что первичная инвазия всегда происходит в ткани корня. У Cytinus и Bdallophyton лучи соматической ткани редко распространяются в ткани побега, хотя их развитие, как и у других Раффлезиевых, протекает интраматрикально. По внешнему облику зрелый спорофит Cytinus более сходен с таковым,, например, Orobanche1 нежели со спорофитом других Раффлезиевых. Для Cytinus типичны хорошо развитые генеративные побеги, несущие мощное соцветие.
Для Rafflesia и, вероятно, также Sapria и Brugmansia (Rhizanthes) характерна полная редукция генеративного побега до цветка, интра- матрикальное развитие и возможное, но не облигатное проникновение соматических лучей в ткани побега растения-хозяина (Schmucker, 1959b, и др.). Развитие этих паразитов происходит анизофазно с развитием их хозяев. I
Для Pilostyles, Apodanthes и Mitrastemon типично облигатное интра- матрикальное распространение соматических лучей паразита из тканей корней в ткани побегов, особенно в область апекса побега-хозяина. Для Pilostyles и Mitrastemon установлен изофазный тип развития паразита и хозяина (Kuijt, 1960, 1969). Для этих растений характерен прямой переход от А- к ^-состоянию протосомы.
Изучение эволюции послесеменного периода развития спорофита паразитных цветковых с первой формой паразитизма позволяет прийти к следующим заключениям: I) в результате прогрессирующей редукции зародышей при переходе растений к чужеядному питанию происходит смещение заключительных процессов эмбриогенеза на послесеменной этап развития спорофита; 2) это смещение обусловлено полным угнетением морфогенетической активности эпикотиля, вследствие чего внесеменное развитие спорофита связано с морфогенетической активностью исключительно базального полюса зародыша; 3) единообразное развитие протосомы из базального полюса зародыша, элиминация эпикотиля и семядолей свидетельствуют о единой форме редукции зародышей у всех растений с первой формой паразитизма; 4) в эволюции протосом отмечено несколько оригинальных направлений, что свитедельствует о различии воздействующих на их эволюцию эктогенетических факторов.
Гомологичность протосом у всех растений с первой формой паразитизма, помимо прямых наблюдений над прорастанием, хорошо подтверждается строением семени в целом. Характерное расположение редукци- рованного зародыша в семени растений, у которых изучалось прорастание (Orobanehe1 Cynomorium1 Striga orobanehoides, Viseum1 Areeuthobium и др.), строго повторяется у всех растений с первой формой паразитизма, у которых было исследовано строение зрелых семян.
Еще по теме ПОСЛЕСЕМЕННОЕ РАЗВИТИЕ И МЕТАМОРФОЗ У OROBAN С HE CREN AT А И CY NOMORIU M SONGARICUM:
- ПОСЛЕСЕМЕННОЕ РАЗВИТИЕ И МЕТАМОРФОЗ У THUNIA MARSCHALLIANA
- ПОСЛЕСЕМЕННОЕ РАЗВИТИЕ И МЕТАМОРФОЗ У ПРЕДСТАВИТЕЛЕЙ СЕМ. PYROLACEAE
- ПОСЛЕСЕМЕННОЕ РАЗВИТИЕ, МЕТАМОРФОЗ И ЭВОЛЮЦИЯ ПРОТОСОМЫ У CUSCUTA
- МЕТАМОРФОЗ
- МЕТАМОРФОЗ, РЕДУКЦИЯ И НЕОТЕНИЯ И СИСТЕМЫ МОДУСОВ ЭВОЛЮЦИОННЫХ СТРУКТУРНЫХ ПРЕОБРАЗОВАНИЙ
- ПРОДОЛЖИТЕЛЬНОСТЬ РАЗВИТИЯ
- 18.7. ЦИКЛ РАЗВИТИЯ ПАРАЗИТОВ И ОРГАНИЗМ ХОЗЯИНА
- Канализация развития
- 6. 4. 7. Развитие биосферы
- Эмбриональное развитие