ВНУТРИСЕМЕЙНОЙ ПЕРИОД РАЗВИТИЯ ЗАРОДЫША
Сем. Pyrolaeeae. Поскольку представители семейства значительно различаются между собой по степени редукции зародышей, они дают благоприятный материал для изучения модусов морфогенетических эволюционных преобразований.
В то же время изученные представители Грушанковых характеризуются высокой степенью общности начальных этапов эмбриогенеза. Для выявления этого единства существенное значение имеют некоторые особенности эмбриогенеза, на которые ранее не было обращено внимания.Впервые эмбриогенез в сем. Грушанковых был исследован Суэжем (Soueges, 1939Ь) на примере Pyrola rotundifolia. По классификации Cy- эжа, развитие зародыша P. rotundifolia должно быть отнесено к III мегар- хетипу, серии Ay второго периода. Во второй период его классификации типов эмбриогенеза относятся все те растения, в развитии зародыша которых базальная клетка проэмбрио не принимает никакого участия. Однако проведенное нами детальное изучение зародыша грушанок и верт- ляниц показало, что Суэж упустил из виду одну весьма существенную подробность эмбриогенеза этих растений. Суэж полагал, что у P. rotundifolia элемент п' происходит от са через cd в результате деления ci. Он считал также, что п' совместно с сЪ образует суспензор s, который находится в области микропилярного гаустория эндосперма и отмирает вместе с гау- сторием, тогда как элемент п, также происходящий от са через ci, в дальнейшем образует со, т. е. медианную часть чехлика корня.
Такое толкование эмбриогенеза Pyrola ошибочно, и ошибка. Cyэжа объясняется весьма интересным явлением, которое было упущено им из виду: вслед за первым делением ядра базальной клетки сЪ одно из дочерних ядер отходит в базальную область вытянувшегося в трубку проэмбрио, а другое — в сторону апикальной клетки (Терёхин, 1962а, 19626). У клеточной перегородки, разделяющей обе дочерние клетки, образуются вакуоли, и создается впечатление, что дочерняя клетка п', примыкающая к апикальному комплексу клеток, происходящему от са, этому же
комплексу и обязана своим происхождением.
В действительности эта клетка (клетка т или d в настоящем исследовании) происходит от сЪ (рис. 27, I).В результате исследования последующих этапов развития зародыша грушанок и вертляниц было выяснено, что элемент пг не только происходит от базальной клетки двуклеточного проэмбрио сЪ, но и занимает в структуре зародыша то положение, которое Суэж приписывал элементу Iг. Таким образом, даже если исходить из системы Суэжа, развитие зародыша грушанок и вертляниц должно быть отнесено не ко второму, а к первому периоду.
Интересно, что Вейе-Бартошевская (Veillet-Bartochewska, 1958, 1959, 1960а), изучавшая эмбриогенез у представителей порядка Ericales1 в одной из последних работ пришла к выводу, что изученная ею Clethra alni- folia по способу развития зародыша должна быть отнесена к первому, а не ко второму периоду системы Суэжа.
Явление перемещения ядер в клетках проэмбрио Грушанковых не только позволило найти ключ к правильной интерпретации эмбриогенеза этих растений, но и установить общую для этой группы растений черту эмбриогенетического развития. ,
Рассмотрим некоторые другие особенности эмбриогенеза Грушанковых. Одной из таких особенностей является, по нашему мнению, значительная вариабельность в поведении разных бластомеров зародыша на средних и поздних этапах эмбриогенеза. Эмбриогенез у относительно менее специализированной Orthilia seeunda и более специализированного Moneses uniflora являет собой переход к таким крайним формам специализации эмбриогенеза, которые представлены Hypopitys1 Allotropa1 Mono- tropa.
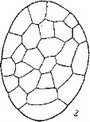
Как можно видеть из рис. 27, I1 зигота О. seeunda после проникновения внутрь эндосперма делится в терминальной области поперечной перегородкой. Каждая из вновь образовавшихся таким образом дочерних клеток также делится поперечно, образуя линейную тетраду бластомеров. Однако вполне сформированный нитевидный проэмбрио состоит часто не из 4, а из 5—6 клеток, расположенных друг над другом. При этом самые верхние этажи клеток образуются в результате дополнительного поперечного деления дочерних клеток от ei (рис.
27, /). Таким образом, в формировании нитевидного проэмбрио обнаруживается некоторая вариабельность, которая усугубляется еще и тем, что в отдельных случаях бластомер еа делится не поперечной, как обычно, а продольной перегородкой, формируя Т-образную тетраду бластомеров.Дальнейшую судьбу бластомеров в этом варианте развития нам проследить не удалось. Обычно же бластомер еа делится поперечно, образуя элементы I ж Г (это соответствует обозначениям ее и ed в нашей работе 1962 г.). Однако обозначение этих бластомеров через 1жУ более правильно, поскольку, как мы уже выяснили, способ развития зародыша этих растений не может быть отнесен ко второму периоду по системе Суэжа. Далее клетки этажей I1 V, т, с или d и / верхней и нижней дочерних клеток от т делятся продольными перегородками, при этом последовательность делений по этажам и положение вертикальных перегородок внутри этажей также варьирует (рис. 27, I). Варьирует и ориентация делений в этажах т и d. Повторными вертикальными делениями в этажах I ж Г образуются верхние и нижние октанты, а за ними следуют периклинальные деления, маркирующие дифференциацию эмбриодермы. Иногда типичные картины дифференциации эмбриодермы в этаже I нарушаются. Антиклинальные деления в клетках эмбриодермы происходят сопряженно со специфическим процессом смещения положения клеток относительно друг друга.
Нетрудно заметить, что нарушения строгой детерминированности развития при дифференциации эмбриодермы, изменение положения клеток созревающего зародыша, отсутствие дифференциации периблемы
alt="" />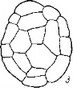
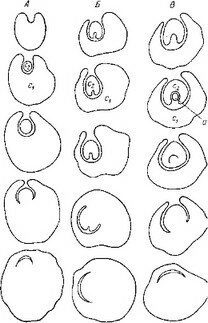

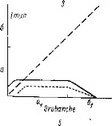
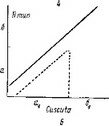
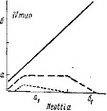
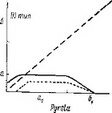
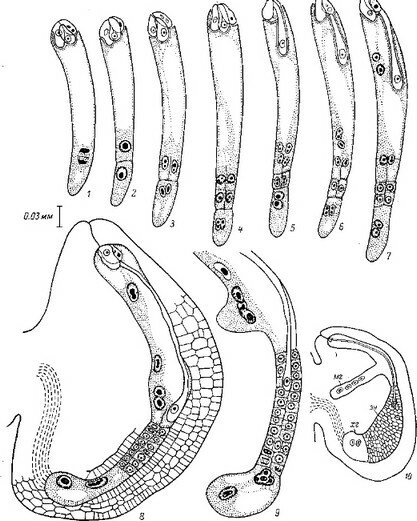
I — Orthilia sccunda; 11 — Moneses uniflora\ III — Hypopitys monotropa\ IV — Monoiropa uniflora\
V — Allotropa virgata.
и плеромы — все эти явления общие и для зародышей Грушанковых, и для зародышей Заразиховых, и, по-видимому, для зародышей всех высокоспециализированных паразитных растений.
По сравнению с эмбриогенезом О. secunda эмбриогенез Moneses uniflora характеризуется следующими особенностями. При общности первых этапов эмбриогенеза у обоих растении на средних этапах развития эмбриогенез М. uniflora отличается значительно более высокой вариабельностью в поведении бластомеров, особенно тех из них, которые произошли от са (рис. 27, II). Морфогенетическая активность клеток апикальной области проэмбрио M. uniflora затухает значительно раньше, чем у О. secunda, еще до дифференциации эмбриодермы.
У исследованных нами наиболее высокоспециализированных представителей семейства — Hypopitys monotropa, Allotropa virgata и Monotropa uniflora — редукция автотрофной организации зародыша зашла наиболее далеко (Терёхин, 1962а, 19626, 1962в, 1962г; Терёхин, Яковлев, . Особенности начальных стадий развития зиготы у H. monotropa, virgata, M. uniflora не оставляют сомнений в принципиальном единстве их эмбриогенеза с эмбриогенезом остальных Грушанковых. В сущности весь эмбриогенез у этих растений сводится к 2—4 делениям, и важна при этом стабильность признаков, характеризующих первое деление (положение и место перегородки, время деления и т. п.), и вариабельность ориентации и последовательности второго и последующих делений (рис. 27, Ш-У).
G формальных позиций классификации Суэжа эмбриогенез Monotropa, Hypopitys и Allotropa должен быть отнесен ко второму периоду его системы, так как в эмбриогенезе этих растений базальная клетка сЪ не принимает участия в построении тела зародыша. Однако было бы ошибкой сделать такое заключение, поскольку имеется достаточно много признаков, не оставляющих сомнения в общности эмбриогенеза этих растений с эмбриогенезом остальных Грушанковых. Сходство эмбриогенетического развития указывает определенным образом на единственно возможный способ эволюционных изменений зародышей — терминальную аббревиацию, осуществляющуюся путем последовательной элиминации конечных этапов развития. Очевидно при этом, что явления редукции сопровождаются ослаблением жесткой детерминации в эмбриогенезе; причем это ослабление проявляется на все более и более ранних этапах эмбриогенеза (Orthilia—Moneses—Hypopitys).
Сем. Gentianaceae. Сведения о развитии зародышей паразитных представителей семейства очень скудны. Известно, что в результате первых делений у Voyria coerulea, V. trinitatis, V. uniflora, V. tenella, Cotylanthera tenuis, Voyriella parviflora и Leiphaimos sp. образуется линейная тетрада бластомеров, а затем апикальные клетки проэмбрио делятся вертикальными перегородками; на этом развитие зародыша заканчивается (Johow, 1885, 1889; Figdor, 1897; Oehler, 1927). Таким образом, наблюдаемая в ряду паразитных видов этого семейства редукция зародыша в зрелом семени происходит на фоне сходства начальных этапов эмбриогенеза. Редукция зародышей и здесь происходит, как видно, путем последовательной терминальной аббревиации.
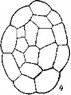
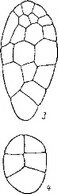
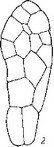
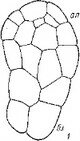
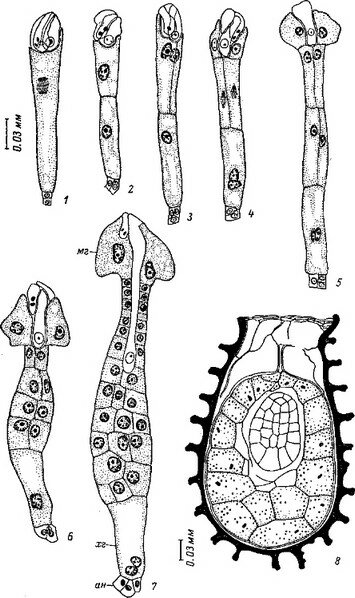
Сем. Burmanniaceae. У исследованных видов рода Burmannia (В. coelestis, В. Candida, В. capitata, В. javanica) и Apteria Se- tacea зигота и базальная клетка двуклеточного проэмбрио делятся поперечными перегородками. Апикальная клетка делится продольно однажды или дважды, и на этом развитие зародыша заканчивается (Treub, 1883b, Johow, 1885, 1889; Ernst, 1909; Ernst, Bernard, 1911—1912). В некоторых случаях (у Apteria setacea) в клетках квадрантов закладываются периклинальные перегородки, означающие дифференциацию эмбриодермы (рис. 28). С другой стороны, эмбриогенез у В. coelestis и Candida может быть завершен на 3-, 4- и 6-клеточной стадии развития
ft ..? Рис. 28. Эмбриогенез некоторых представителей сем. Burmanniaceae.
I Burmannia disticha; 2 — Apteria setacea.
(Ernst, 1909; Schmid, 1917; наши наблюдения над эмбриогенезом В. disticha). Вариабельность конечных этапов эмбриогенеза при стабильном характере первых делений у Бурманниевых — еще одно очевидное свидетельство редукции посредством последовательной элиминации конечных стадий развития.
Данные по эмбриогенезу рода Thismia весьма противоречивы (Meyer, 1909; Ernst, Bernard, 1911; Schmid, 1917, Pfeiffer, 1918; Goebel, Sues- senguth, 1924).
ЗАКЛЮЧЕНИЕ
* Изучение аллелопаразитных растений с третьим способом чужеяд- яости позволяет прийти к следующим заключениям: структурные, эволюционные преобразования зародышей аллело- паразитов характеризуются явлениями редукции и субституции; 2) общая редукция зародышей тем более выражена, чем более специализирован в направлении паразитизма тот или другой вид аллелопаразитного растения; 3) форма дифференциальной редукции соответствует способу перехода проростков спорофита к чужеядному питанию, у зародышей полностью элиминируются семядоли и эпикотиль и происходит гистогенная редукция радикулы при сохранении морфогенетического потенциала в базальной области проэмбрио; 4) для всех изученных видов характерно послесеменное развитие с метаморфозом; 5) форма метаморфоза коррелирует со способом перехода растений к чужеядности; 6) основной модус осуществления структурных эволюционных преобразований зародыша — последовательные терминальные преобразования.
Еще по теме ВНУТРИСЕМЕЙНОЙ ПЕРИОД РАЗВИТИЯ ЗАРОДЫША:
- ИЗУЧЕНИЕ ВНУТРИСЕМЕЙНОГО периода РАЗВИТИЯ ЗАРОДЫША
- ИЗУЧЕНИЕ ВНУТРИСЕМЕЙНОГО ПЕРИОДА ЭМБРИОГЕНЕЗА
- РАЗВИТИЕ ЗАРОДЫША И ПЛОДА
- РАЗВИТИЕ ЗАРОДЫША И ПЛОДА
- 2-5. Развитие зародышей и эволюция. Жоффруа Сент-Илер
- ГЛАВА IV НАЧАЛЬНЫЙ ПЕРИОД РАЗВИТИЯ ЖИЗНИ
- Развитие экологии в последарвиновский период.
- ОБРАЗОВАНИЕ И РАЗВИТИЕ БОЛОТ ОТ ГОЛОЦЕНАДО НАСТОЯЩЕГО ПЕРИОДА
- ГЛАВА 1 Представления о развитии живой природы в додарвиновском период
- ТРАНСПЛАНТАЦИЯ ЗАРОДЫШЕЙ
- ТРАНСПЛАНТАЦИЯ ЗАРОДЫШЕЙ
- ЗАРОДЫШИ В ЗРЕЛЫХ СЕМЕНАХ
- ОЦЕНКА, КУЛЬТИВИРОВАНИЕ И ХРАНЕНИЕ ЗАРОДЫШЕЙ
- ОЦЕНКА, КУЛЬТИВИРОВАНИЕ И ХРАНЕНИЕ ЗАРОДЫШЕЙ