Возникновение и эволюция рифовых сообществ
История[‡‡‡‡‡‡] рифовых сообществ весьма хорошо изучена благодаря тому, что поры рифовых построек — одно из основных вместилищ нефти. Биогенные известковые постройки, аналогичные современным коралловым рифам, известны с докембрия.
В то время их образовывали прокариоты. Создаваемые ими постройки (строматолиты) имели толщину до 30 м и длину до нескольких сотен метров. В современных морях строматолиты в кон-
Рис. 3.63 Современные строматолиты. Общий вид и микроскопическое строение. По Littler, Littler (2001).
це XX века были найдены у Багамских островов (рис. 3.63). В раннем кембрии (до позднего кембрия) появляются рифы, впервые образованные скелетами многоклеточных организмов — археоциатами — вымершим таксоном, сходным с или принадлежащим к губкам (рис. 3.64). Затем до среднего ордовика рифы отсутствовали.
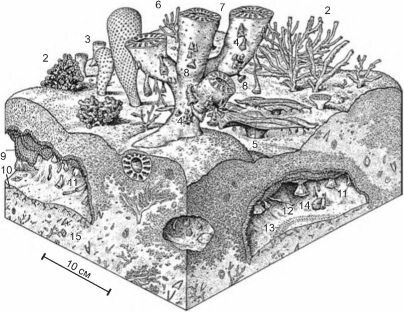
Второй раз рифовые сообщества многоклеточных возникли в середине ордовика и существовали до силура - среднего девона. В период максимального развития рифы строили губки, кораллы (представленные ныне вымершими таксонами: табулятами, ругозами и геликоидеями), мшанки, цистоидеи (вымерший таксон иглокожих) и некоторые другие,
Рис. 3.64. Реконструкция нижнекембрийского рифа.
1 — Renalcis (известковая цианобактерия), 2, 3, 4, 5 — археоциаты, 6, 7 — губки (?), 8 — критические кораллоподобные организмы; 9 — начинающий формироваться волокнистый цементирующий осадок внутри пустот; 10 — ходы в грунте груптоедов: 11 — критические археоциаты и кораллоподобные организмы; 12 — трубки неясного происхождения; 13 — следы трилобита; 14 — ботриодиды; 15 — осадок с остатками скелетов.
По Wood (1998).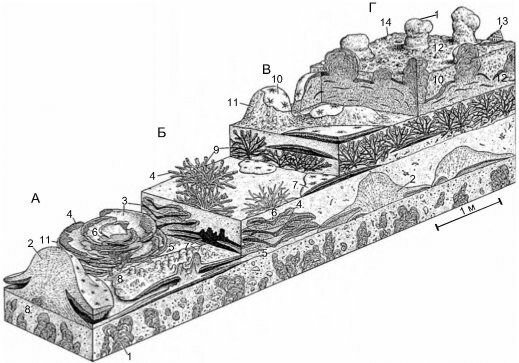
Рис. 3.65. Реконструкция позднедевонского рифа (360 млн. лет назад).
В осадках рифа можно выявить отчётливые стадии, связанные с уменьшением глубины. Эти стадии интерпретируют как четыре сообщества, сменяющих друг друга: А — начальная стадия карбонатного осадкообразования, возможно инициированная увеличением глубины, для неё характерно колонии больших строматопорид на неподвижном грубообломочном осадке: купольных (Actinostroma sp.), завитых листовидных (f Actinostroma sp.) и плоско-столбчатых. Эти строматопориды сильно изъедены Renalcis и мелкими организмами, особенно под поверхностью грунта. Б — следующая зона, характеризующаяся зарослями ветвистых строматопорид (Stachyocles sp.), и тонкими, пластинчатыми строматопоридами возвышающимися над осадком. В — зона экстенсивного роста больших холмов микробиалитов, которое источены строматопоридами (?ClathrocoПопа spissa). Г — столбчатые головы строматолитов, развивающиеся на крупнопесчаном мелководье, с пятнами больших онколитов (вероятно образованных микробионтами) и большими гастроподами.
1 — строматолиты; 2 — купольная (domal) строматопорида (Actinostroma); 3 — inferred завитые листовидные - строматопориды (p. Actinostroma sp.); 4 — известковые цианобактерии (Renalcis); 5 — волокнистый цементирующий осадок; 6 — geopetal sediment infill; 7 — пластинчатые строматопориды; 8 — морские лилии; 9 — ветвистые строматопориды (Stachyocles sp.); 10 — инкрустирующие строматопориды (?ClathrocoiIona spissa); 11 — ми- кробиалиты; 12 — грубообломочный осадок; 13 — гастроподы; 14 — онколиты.. Canning Basin, западная Австралия. По Wood (1998).
ныне вымершие таксоны (рис. 3.65). Первые кишечнополостные с известковым скелетом (Tabulata) появились в ордовике. Затем появляется вторая группа кишечнополостных с известковым скелетом — ругозы. С появлением ругоз рифовые постройки достигли современных размеров: площадь — сотен квадратных километров, толщина — 1,5 км.
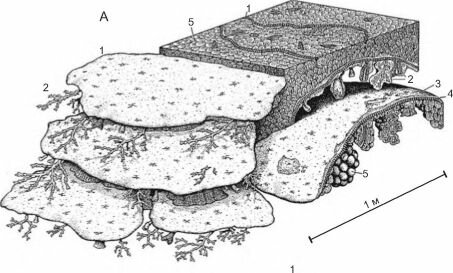
В раннем карбоне вновь возникли органогенные постройки. Однако, в целом, в позднем палеозое отсутствовали организмы с быстрым ростом, способные создать каркас рифа. Поэтому позднепалеозойские органогенные постройки являлись скорее холмами, нежели рифами (рис. 3.66). В их постройке основную роль играли известковые водоросли, а роль животных была подчинённой.
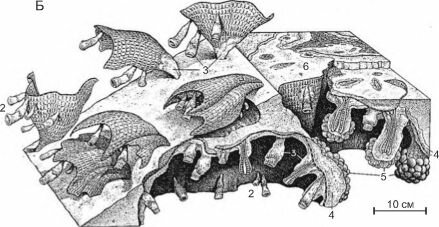
Рис. 3. 66. Реконструкция рифов поздней перми (260 млн. лет назад) .
А. 1,2 — губки (1 — Gigantospongia discoforma); 3 — инкрустирующая водоросль Archaeolithoporella; 4 — отложения микробионтов; 5 —ботриодные выросты (в форме мешочков с узкой шейкой) неясного происхождения. Б. 1 — мшанки (Polypora sp.; Goniopora sp.); 2 — губки; 3 — инкрустирующая водоросль Archaeolithoporella; 4 — отложения микробионтов; 5 — ботриодные выросты; 6 — осадок. The Capitan Reef, Техас и Нью-Мексико. По Wood (1998).
Фактически третий период существования биогенных рифов начался со среднего триаса. До этого, в раннем триасе, рифовые постройки по-видимому отсутствовали. Сначала основную роль в образовании рифов играли водоросли и губки, реже полихеты с известковыми трубками и известковые мшанки, но в середине триаса возникли склеракти- нии, по-видимому из ругоз, утративших ранее известковый скелет, из-за чего их остатки не сохранились, в триасе они вновь приобрели способность к его постройке (Келлер, Ко- смынин, 1997). Постепенно, к позднему триасу, ведущая роль перешла к склерактиниям. В конце триаса наступает новый перерыв в рифообразовании. В этот раз он был почему- то очень краток, на порядок короче других, причём восстановление рифовых сообществ произошло также быстрее, чем в остальных случаях. В самом конце триаса рифы вновь получили развитие. Сначала их основными строителями были склерактинии и водоросли.
В мелу на мелководье ведущая роль в строительстве каркасов рифов переходит к рудис- там (рис. 3.67) — вымершей ныне группе двустворчатых моллюсков, ограниченной в сво-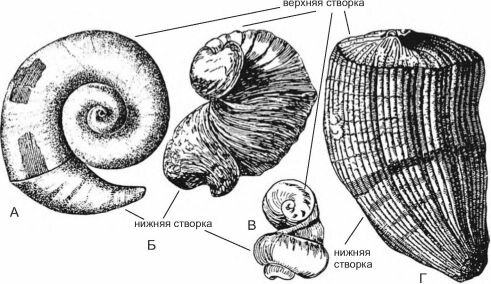
Рис. 3.67. Рудисты.
А — Caprina adversa; Б —Diceras; В —Requienia; Г —Hippurites. По Друщицу (1974) и Бобковой, Пчелинцеву (1960).
Рудисты были одиночными постоянно прикреплёнными к субстрату двустворчатыми моллюсками с неравностворчатой раковиной размером до 2,0 м при 0,6 м в диаметре, со стенками толщиной до 10 см. Возникнув в поздней юре, в мелу они дали вспышку таксонообразования (известно около 1000 видов), но полностью вымерли к началу кайнозоя. Позднеюрские и раннемеловые рудисты имели преимущественно спирально закрученные створки, к субстрату прикреплялась или маленькая створка, или большая. Аналогом рудистов являются современные Chamidae (к прикреплённому образу жизни перешли недавно и не имеют симбионтов) и устрицы Ostre- idae. Позднемеловые рудисты были преимущественно конические с маленькой конической створкой. Рудисты не были так чуствительны к условиям среды, как кораллы. Обитали исключительно в тропиках, где образовывали рифы, не уступавшие по масштабности рифам склерактиний.
ём распространении Тетисом. Рудисты обитали и на больших глубинах, но не создавали там рифов.
Поскольку рудисты — одиночные организмы, а не колониальные, как склерактинии, образованные ими рифы скорее имели вид зарослей, нежели коралловых рифов. «В течение мелового времени разнообразие рудистов возрастало за счёт развития новых адаптивных типов (пряморастущих, планоспирально завитых, лежавших свободно на субстрате, устрицеподобных и пр.); возникали скульптурированные формы, ребра и гребни на раковинах которых способствовали прочному соединению прямостоящих форм; некоторые формы цементируются к соседним особям. У многих форм верхняя створка становится прободенной или покрывается мантийной тканью, что облегчало симбиотическое отношение с зооксантеллами» (Невесская, 1999: 207).
Глубже склерактинии сохраняли свою ведущую роль. К концу мела рудисты вымерли и основными рифостроителями вновь стали склерактинии. Но это продолжалось недолго.В конце мела - палеоцене происходит очередной перерыв в рифообразовании. На отмелях и мелководьях карбонатных платформ, где в меловое время были развиты рифовые структуры, в первой половине палеоцена они исчезли, затем начали формироваться небольшие постройки в основном из кораллиновых водорослей и лишь местами — из губок и склерактиний (Bryan, 1991), и только в олигоцене вновь возникли сложно построенные коралловые рифы, каркасостроителями которых были кораллы, кораллиновые и зеленые водоросли, мшанки и некоторые фораминиферы, двустворки и гастроподы, но постепенно ведущая роль в них опять переходит к склерактиниям. Современный облик коралловые рифы приобретают в плиоцене - миоцене. Тропическая флора известковых красных водорослей (и, очевидно, растительность) существенно изменилась в начале неогена (Перестенко, 1994), т. е. одновременно с началом формирования коралловых рифов. На рис. 3.22 показан средний возраст современных родов Scleractinia. Однако, что касается родов рифообразующих Scleractinia, то они возникли не ранее олигоцена, причём обитающие в Атлантике рода отнюдь не старше Индо-Пацифических, как это может показаться из этого рисунка. По-видимому, коралловые рифы возникли в Тетисе, затем, после разделения Тетиса произошло независимое образование родов и видов в Атлантике и Индо- Пацифике, приведшее к современной биогеографической картине.
Таким образом, рифовые сообщества формировались в течение фанерозоя неоднократно. Многие рифостроители — фильтраторы эпифауны. Но, наиболее крупные рифовые постройки возводят и возводили фотосинтетики: известковые водоросли (кембрийские строматолиты, позднее — красные и зелёные водоросли) или фотосимбиотрофные животные, впервые появившиеся, по-видимому, уже в кембрии: археоциаты в кембрии, строматопороидеи (средний ордовик - пермь), табуляты (ордовик - девон), табуляты, некоторые брахиоподы в перми, фузулиниды в карбоне и перми, рудисты и ряд других дву- створок (?пермь,?силур, поздняя юра - ныне) и склерактинии (средний триас - ныне).
Существенную роль в заполнении полостей в рифе играли также обладавшие симбионтами фораминиферы-фузулиниды (карбон - пермь), а в современных рифах — известковые водоросли. Т. е. основой функционирования мелководных рифовых сообществ всегда был фотосинтез.Смена ведущих групп рифостроителей на протяжении фанерозоя носит закономерный характер. Всегда на ранних стадиях формирования и эволюции рифовых сообществ ведущую роль играли известковые водоросли. В раннем палеозое существенную роль имели сине-зелёные водоросли, но позднее их роль становилась всё меньше и с середины мела они уже практически исчезли из морских отложений, с тех пор они образовывали органогенные постройки только в озёрах и солёных маршах аридной зоны. Их место заняли красные и, в меньшей степени, зелёные водоросли. С течением времени, по мере роста разнообразия ведущая роль переходит к рифообразующим животным. В кембрии это были только губки (археоциаты), они играли существенную роль и позднее, особенно в периоды становления рифовых сообществ после перерывов и на глубинах за пределами фо- тической зоны, где их роль существенна и в настоящее время, но в мелководных рифах в процессе их эволюционного развития губок всегда сменяли другие группы. По мере роста биоразнообразия на роли рифостроителей пробуются Brachiopoda, мшанки и многие другие, в том числе большое число ныне вымерших таксонов. Но эффективнее всего эта роль удалась кишечнополостным, разные группы которых доминировали среди рифостроителей с середины палеозоя до настоящего времени. Таким образом, имеется достаточно чёткая и многократно повторявшая схема возникновения и эволюции в геологическом времени рифовых сообществ:
красные известковые водоросли —» губки —» кишечнополостные
Эта же последовательность сообществ наблюдается ныне и при восстановлении коралловых рифов после их разрушения, например, после землетрясений и цунами. Изучение современных сукцессий показывает, что стадии красных известковых водорослей предшествует стадия бурых (фукоидов). Весьма вероятно, что в геологическом прошлом, в периоды, когда отсутствовали рифы, их место было занято аналогичными сообществами макроводорослей, но из-за отсутствия у них каких-либо частей, которые могли бы сохраниться в отложениях, следов их не обнаружено.
Лишь единожды — в мелу — кишечнополостные были вытеснены с первых ролей рудистами. Но, в общем, в истории биосферы с ростом разнообразия и роли кишечнополостных падает роль других рифостроящих групп. Это наблюдается не только во времени, но и в пространстве. Так, начиная с плиоцена и по настоящее время, велика роль водорослей в рифах Атлантики. Это, возможно, объясняется тем, что склерактинии обитают преимущественно в Тихом и Индийском океанах (там их фауна по числу видов на порядок богаче). То же наблюдается и по глубине. Кишечнополостные доминировали и доминируют на малых глубинах, где интенсивность фотосинтеза достаточно велика. Глубже, на внешней кромке шельфа и верхней части склона, фотосинтез неэффективен. Здесь рифы образовывали всегда преимущественно губки, лишь начиная с мела существенную роль местами стали играть склерактинии (подробнее глубоководные рифы рассмотрены в этом разделе ниже).
Почему рифовые сообщества время от времени исчезали на Земле неизвестно. Общепринято считать, что в эти периоды условия жизни на Земле были для них неподходящи (Sheehan, 1985), но это не объяснение, а лишь констатация факта иными словами. Неоднократное исчезновение на Земле экосистем рифов не могло не влиять на общий уровень биоразнообразия биосферы. По последним оценкам до 1 миллиона видов животных и растений (в том числе четверть всех морских рыб) связано с тепловодными коралловыми рифами. Если бы на Земле исчезли коралловые рифы, существенная часть этих видов вымерла бы. Нет оснований полагать, что исчезновение рифов в предшествующие геологические эпохи имело менее катастрофичные последствия для морской биоты.
Основания современных рифовых построек начали формироваться в конце мезозоя - начале кайнозоя. Рост рифа компенсировал увеличение глубины вершины основания рифа из-за трансгрессий или тектонических движений. Сформировавшиеся в результате известковые массивы достигают толщины 2 км и более. В периоды регрессий и положительных тектонических движений рост рифов происходил по периметру платформы, вершина же платформы, если она возвышалась над поверхностью, подвергалась эрозии. Последний из таких периодов был во время максимального оледенения 130-150 тыс. лет назад. В результате, все современные коралловые рифы представляют собой тонкую плёнку (толщиной 3-33 м) на поверхности плейстоценовых платформ, эродированных во время последнего ледникового периода.
Еще по теме Возникновение и эволюция рифовых сообществ:
- Рифовые сообщества Особенности рифовых сообществ
- Возникновение теории эволюции
- 14. Эволюция организмов и эволюция сообществ
- ЭВОЛЮЦИЯ ПРОТОБИОНТОВ И ВОЗНИКНОВЕНИЕ ПЕРВИЧНЫХ ОРГАНИЗМОВ
- ГЛАВА 7 Возникновение учения о микроэволюции. Популяция — элементарная единица эволюци
- Глава 13 Эволюция сообществ насекомых
- Продуктивность сообществ и жизненные стратегии водорослей. Морфофункциональный подход к изучению сообществ макрофитов
- РАЗРУШИТЕЛИ КОРАЛЛОВЫХ РИФОВ
- СОСТАВ СООБЩЕСТВ И КОМПЛЕКСЫ СООБЩЕСТВ
- Рифовый олень Odocoileus virginianus clavium Barbour et G. Allen,1922 (I, 428)
- 1.6. ВОЗНИКНОВЕНИЕ МНОГОКЛЕТОЧНОСТИ
- 11-2* Возникновение нового