ОЛИГОТРОФНЫЕ МИКРООРГАНИЗМЫ
Олигонитрофилы были изучены многими исследователями (Kauffman, 1951; Мишустина, 1955; Паринкина, 1960; Мальцева, 1968; Клевенская, 1971, и др.).
И. Е. Мишустина, исследовавшая большую коллекцию оли- гонитрофильных микроорганизмов, пришла к выводу, что на МПА развивается не более 30—40% этих микроорганизмов. При анализе почвы на бедной азотом среде весьма нередко учитывается значительно больше микроорганизмов, чем на МПА. Наблюдаемая разница должна быть отнесена за счет олигонитро- Филов, не растущих или плохо растущих на МПА.
Олиготрофные микроорганизмы вообще и олигонитрофилы в частности должны составлять существенную группировку ъ почвах, в которые поступает свежее, бедное азотом органическое вещество, т. е. там, где происходит образование продуктов распада растительных остатков с низким отношением C/N. Можно ожидать, что в длительно парующих почвах их численность будет редуцироваться. Это вполне подтверждается данными Мишустиной, представленными на рис. 16. Из них видно, что в паровавшей около 50 лет дерново-подзолистой почве Сельскохозяйственной академии им. К. А. Тимирязева на среде Эшби (содержащей незначительное количество азота) учитывалось практически столько же бактерий, сколько на МПА. В почве, занятой бессменной рожью, численность олигонитрофильной груп-
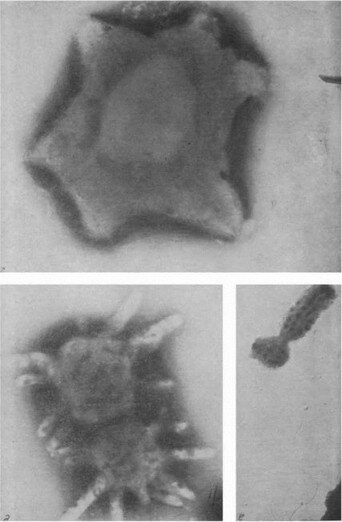
Рис. 15. Развитие разных бактерий в среде, содержащей 0,5% сахара 1 — Ps. fluorescens, 2— Microcyclus major
пировки резко возросла и существенно превышала число бактерий, учитываемых на МПА.
И. Л. Клевенская (1971), изучавшая наличие олигонитрофи- лов в различных почвах Сибири, нашла, что в отдельных случаях их относительная численность сильно колеблется. Иногда их число равно показателям, получаемым на МПА, а нередко в 5— 12 раз превышает численность банальных сапрофитов. Несомненно, это связывается с поступлением в почву растительных остатков. В среднем отношение олигонитрофилов к типичным сапрофитам колеблется между 2—4. Клевенская отмечает, что коррелятивная связь наибольшей силы обнаруживается между численностью олигонитрофилов и содержанием в почве углерода.
В зональных типах почв равнинной территории наблюдает* ся закономерное увеличение олигокарбофилов от подзолов и подзолистых почв к дерново-подзолистым и серым. Их числа достигает максимума в выщелоченных черноземах и резко снижается в южных черноземах и каштановых почвах. Относитель*
ная численность олигонитрофилов в микробных ценозах доходит до максимума в подзолах и подзолистых почвах. Это свидетельствует о большой роли олигонитрофилов в генезисе почв подзолистого ряда.
Среди олигонитрофилов имеются бактерии, принадлежащие к разным систематическим группам, и актиномицеты. В южных
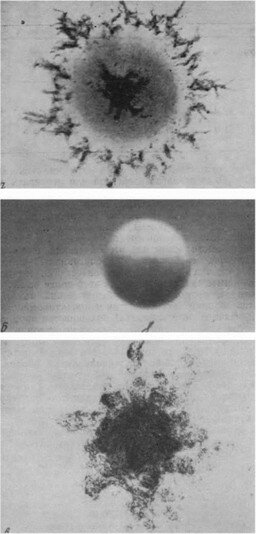
Рис. 16. Сезонные изменения численности микроорганизмов в дерново-подзолистой почве (по И. Е. Мишустиной)
а — бессменный пар, б — бессменная озимая рожь, 1 — МПА, 2 — среда Эшби
почвах относительное значение актиномицетов возрастает. Многие олигонитрофилы могут связывать небольшие количества молекулярного азота.
Многочисленны в почве и олигокарбофильные микроорганизмы, интенсивное изучение многих форм которых происходит в настоящее время. Общеизвестно, что имеются микроорганизмы, хорошо размножающиеся в дистиллированной воде, используя крайне небольшой запас имеющихся там органических соединений. В. И. Романенко и Е. П. Никифорова (1974) показали, что фильтрованная речная вода, почти полностью освобожденная от бактериальных клеток и взвешенных минеральных и органических частиц, является хорошей средой для размножения некоторых микробов. При 20—26° на такой среде число бактерий удваивается через 1—3 часа.
Для изолирования олигокарбофилов используют бедные среды— слабо агаризованные фульваты, агаризованную почву, минеральные среды с низким содержанием органических соединений и т. д. Некоторые олигокарбофилы ассимилируют метиловый спирт и монокарбоновые кислоты. Для них, очевидно, характерно включение в обмен значительных количеств С02 (Доман и др., 1965).
Обзор интересных форм почвенных микроорганизмов, выделенных на специфических средах, дан в статье Д. Н. Никитина «Новые формы микроорганизмов» (1971), к которой мы отсылаем желающих более подробно ознакомиться с недавно выделенными из почвы микробами. Здесь же мы делаем лишь краткий обзор наиболее интересных их видов.
К группе олиготрофных организмов относятся бактерии с необычной морфологией, являющиеся представителями семейства Spirillaceae. Это слабо или более сильно изогнутые палочки с жесткой клеточной стенкой.
Микроорганизмы отмеченной группы были изучены рядом исследователей (0rskov, 1928; Громов, 1963; Claus et al., 1968; Никитин, 1971, и др.).
На бедных средах обнаруживается развитие почкующихся бактерий, относимых к семейству Hyphomicrobiaceae. В эту группу следует отнести представителей рода Pedomicrobium, разлагающих комплексные соединения фульвокислот с полуторными окислами. Типичные почкующиеся бактерии имеют прото- плазматические выросты — «гифы», на которых образуются почки, являющиеся органами размножения.
Особую группу составляют также бактерии, образующие широкие выросты «простеки», образованные клеточной стенкой и цитоплазматической мембраной одновременно. Описаны два рода подобных микробов: Prosthecomicrobium, образующий заостренные простеки, и Ancalomicrobium, имеющий длинные округлые выросты. Эти бактерии размножаются почкованием.
Л. В. Васильевой (1972а) из почвы выделена культура Prosthecomicrobium, близкая к ранее описанному виду Pr. enhydrum. Была изолирована также культура Ancalomicrobium, обладающая ветвящимися, длинными простеками. В отличие от первой культуры она не обладала подвижностью.
К почкующимся бактериям близки представители семейства Caulobacteriaceae, которые широко распространены во многих почвах. Клетки бактерий этой группы имеют вибриоидные или бактериоидные клетки, снабженные стебельком. Стебелек является частью бактериальной клетки — ее отростком. Он вместе с клеткой окружен общей оболочкой. В молодом возрасте клетки
Рис. 17. «Новые» формы микроорганизмов под электронным микроскопом (а—в — по Д. И. Никитину; г—е — по Л. В. Васильевой)
а — Renobacter vacuolatum, б — Tuberoidobacter mutans, в — Ancalomicrobium sp., г — Stella humosa, d — Prosthecomicrobium, e — Agrobacterium sp.
 />Рис. 17 (продолжение)
/>Рис. 17 (продолжение)подвижны и обладают одним жгутиком. Клетки, образовавшие стебелек, теряют подвижность. Стебелек вырастает на месте прикрепления жгутика. Он служит для прикрепления к субстрату. В чистых культурах, где плотность популяции высокая, многие клетки прикрепляются друг к другу, давая розетки. На конце стебельков имеется аморфное внеклеточное вещество, способствующее скреплению клеток.
К настоящему времени описано значительное число видов каулобактерий. Они аэробы. Их чистые культуры не развиваются на МПА, но дают хороший рост, если компоненты питательной среды разводятся в 50—100 раз. Каулобактерии могут усваивать многие органические соединения, если они находятся в низких концентрациях. Бактерии рода Caulobacter до 12% углерода биомассы используют из углекислоты. Большинство чистых культур Caulobacter нуждается в факторах роста (витамины В2, В12 и т. д.).
Размножение Caulobacter происходит путем деления перетяжкой.
Представители семейств Hyphomicrobiaceae и Caulobacteria- сеае изучались многими исследователями (Перфильев, Габе, 1961; Заварзин, 1961; Аристовская, 1965; Stanley, 1968; Беляев,. 1970; Никитин, 1971; Васильева, 1972, и др.).
Многие бактерии, представители семейства Bacteriaceae, также являются олиготрофами. Сюда следует отнести весьма оригинальную по морфологии бактерию Seliberia stellata, выделенную Т. В. Аристовской (1965). Клетки этого гетеротрофного микроба откладывают гидроокись железа как побочный продукт минерализации фульвокислоты. Длинные палочковидные клетки микроба спирально закручены и соединены в звездообразные комплексы (розетки), содержащие часто несколько десятков клеток. На обычных питательных средах Seliberia не растет. Микроб был выделен с бедной среды, содержащей фульваты. Он размножается делением.
С бедной же питательной среды была выделена своеобразная бактерия Agrobacterium polysphaeroides (Никитин, Васильева, 1968), клетки которой имеют многочисленные выросты, образуемые клеточной стенкой. Между цитоплазматической мембраной и клеточной стенкой у этой бактерии имеется широкий слой, очевидно, мукополипептидной природы. Микроб является пери- трихом.
Д. И. Никитиным на бедной среде был изолирован из почвы Tuberoidobacter vacuolatum — бактерия, близкая к предыдущему виду, однако имеющая ряд существенных отличий. Клетки Tuberoidobacter’a более крупные, обычно имеют перетяжку, делящую клетку как бы на две части. Выросты клетки существенно длиннее, чем у Agrobacterium, и беспорядочно расположены на поверхности клетки. У Agrobacterium они расположены по спирали.
Оба отмеченных вида размножаются неравномерным делением.
С бедной питательной среды Васильева (1972а) выделила интересную по ряду свойств бактерию—Stella humosa, систематическое положение которой пока не установлено. Организм имеет форму морской звезды с радиально лучевой симметрией. Клетки подвижны (монотрихи) и размножаются делением, причем плоскость деления перпендикулярна плоскости клетки. Бактерия растет только на сильно разбавленных лабораторных средах. Обладает весьма слабой ферментативной активностью.
Ограничивая список описанными организмами, мы можем отметить, что олиготрофы весьма многочисленны в почве и разнообразны по систематическому положению. Их физиологические свойства также нетождественны. Некоторые из них, как Stella humosa и Prosthecomicrobium, не растут даже на простых углеводах и на белках. Другие, как Tuberoidobacter, более активны и ассимилируют даже некоторые циклические соединения. Отмеченные моменты позволяют думать, что отдельные виды олиготрофов участвуют на разных этапах переработки органических соединений и неодинаково богато представлены в разных почвах. Это мнение могут подтвердить лишь будущие исследования.
Микрофотографии некоторых олиготрофных бактерий даны t\d рис. 17.
М
Еще по теме ОЛИГОТРОФНЫЕ МИКРООРГАНИЗМЫ:
- Биогеоценозы олиготрофного типа
- ВЛИЯНИЕ ИЗМЕНЕНИЙ ВОДНОГО РЕЖИМА ОЛИГОТРОФНЫХ БОЛОТ НА ИХ ЭКОСИСТЕМЫ К. Д. Романюк
- КОНКУРЕНЦИЯ ОЛИГОТРОФНОГО И ЕВТРОФНОГОЗАБОЛАЧИВАНИЯ В ТАЕЖНОМ ЛАНДШАФТЕ С БЛОКОВОЙ СТРУКТУРОЙ
- ХИМИЧЕСКИЙ СОСТАВ, ВЫНОС МИНЕРАЛЬНЫХ И ОРГАНИЧЕСКИХ КОМПОНЕНТОВ ВОДАМИ ОЛИГОТРОФНЫХ БОЛОТНЫХ МАССИВОВ
- ВЛИЯНИЕ ОСУШИТЕЛЬНОЙ МЕЛИОРАЦИИ НА БИОПРОДУКТИВНОСТЬ СОСНОВЫХ ОЛИГОТРОФНЫХ БОЛОТСЕВЕРА БЕЛАРУСИ
- 3. Ферменты микроорганизмов.
- Концепция ненасыщенностикомплекса почвенных микроорганизмов
- ЭПИФИТНЫЕ МИКРООРГАНИЗМЫ
- Физиология и биохимия микроорганизмов
- РАЗВИТИЕ МИКРООРГАНИЗМОВ В ПЛЕНКАХ И КАПИЛЛЯРАХ
- ПРИРОДА ЯВЛЕНИЯ АДГЕЗИИ МИКРООРГАНИЗМОВ
- ВЫЯВЛЕНИЕ МИКРООРГАНИЗМОВ, УЧАСТВУЮЩИХ В КРУГОВОРОТЕ УГЛЕРОДА
- МИКРООРГАНИЗМЫ РИЗОСФЕРЫ И РИЗОПЛАНЫ
- ОСОБЕННОСТИ ОБМЕНА ВЕЩЕСТВ У МИКРООРГАНИЗМОВ
- 2. Биосинтез аминокислот почвенными микроорганизмами.
- Биотермический процесс и термофильные микроорганизмы
- Растворение (лизис) микроорганизмов бактериофагами
- Методы получения чистых культур и культивированияпочвенных микроорганизмов
- СИМБИОТИЧЕСКИЕ МИКРООРГАНИЗМЫ АЗОТФИКСАТОРЫ
- СПЕЦИФИКА ПОЧВЫ КАК СРЕДЫ ОБИТАНИЯ МИКРООРГАНИЗМОВ