Примеры формирования ценотических систем
Выше, в разделе «Причины и скорость образования новых видов и экосистем» уже были разобраны некоторые примеры формирования ЦС: островных ЦС, где дефицит деревьев заполняли таксоны семейств с преобладанием трав; Капской и других флор районов со средиземноморским климатом; в Арале после резкого увеличения там солёности; в крупных озёрах; в водоёмах Паратетиса, проходивших периоды распреснения и осолонення (некоторые другие аспекты этой биоты будут рассмотрены ниже в разделе «Биогеография моря») и т.
д. Но во всех этих случаях мы отслеживали образование каких-либо отдельных частей ЦС, лишь ЦС Арала была рассмотрена сравнительно полно. Рассмотрим ещё один пример: образование ЦС пастбищного типа из ЦС консортного типа, пытаясь проследить одновременно судьбу нескольких блоков.Формирование травяного биома
В качестве примера возникновения ЦС путём преобразования рассмотрим возникновение травяных ЦС (ТЦС) — степей, прерий, пампасов, саванн и др. Их формирование обычно связывают с дефицитом влаги (Мордкович, 1982). С.М. Разумовский (1999), проанализировав данные по связи распределения растительности с температурой и влажностью в 531 точках Земного шара, обнаружил, что распределение климаксных фитоценозов не связано с влажностью, что он объяснил способностью сукцессионных систем эффективно регулировать влажность своих биотопов.
С другой стороны, палеонтологические данные свидетельствуют об отсутствии безлесных ЦС в раннем палеогене (палеоцене и начале эоцена). В это время климаксными ассоциациями повсеместно являлись лесные, несмотря на то, что на Земле, несомненно, и в это время были районы с теми же условиями, в которых ныне существуют травяные ЦС.
Эти и другие факты заставляют искать иные причины возникновения и широкого распространения безлесных ЦС, наиболее аргументированной является теория В.В. Же- рихина (1993, 1994, 2003), основные положения которой изложены ниже.
Имеющиеся палеонтологические данные свидетельствуют о том, что ТЦС формировались неоднократно и независимо, но всегда сходным образом. Сначала появлялись крупные листоядные позвоночные, которые замедляли прохождение сукцессий в местах, где появление лесных сукцессионных стадий тормозилось климатически, в первую очередь, из-за снижения количества осадков. В результате площадь, занятая травяными сообществами, увеличивалась. Это, в свою очередь вызывало появление разнообразной травоядной фауны, ещё более тормозившей прохождение сукцессий. Процесс, таким образом, развивался с положительной обратной связью. В конечном итоге формировались эндогенно стабилизированные травяные сообщества с высоким уровнем диффузных коадаптаци- онных связей, весьма лабильных и обеспечивающих гибкое регулирование функционирования ассоциаций в целом в условиях изменчивой среды (т. е. ЦС пастбищного типа). В этих сообществах развивались специфичные для них блоки копрофагов, травоядных насекомых (прямокрылых, термитов), сапрофагов, некрофагов и др. Доминирующими продуцентами стали травянистые растения, адаптированные к быстрому росту после стравливания. Везде для ТЦС характерны очень высокая продуктивность и гораздо более плодородные почвы, чем в соседних лесных ЦС.
Когда впервые возникли ТЦС неизвестно. Вполне можно допустить их формирование на основе папортникообразных. Первые достоверно известные ТЦС отмечены в позднем мелу. Их эдификаторами были растительноядные динозавры (в первую очередь цера- топсиды). Травяной блок был образован не злаками, как ныне, поэтому их нельзя рассма-
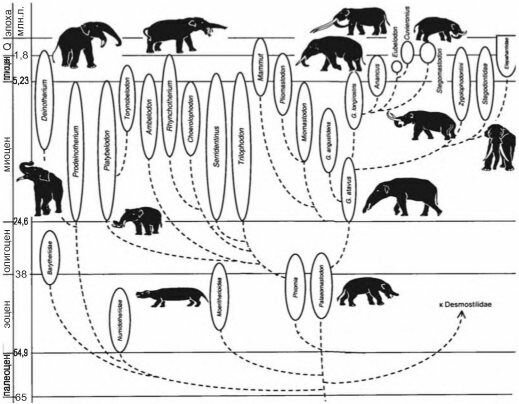
Рис. 4.41 Филогенетическое древо хоботных. По А.К. Агаджаняну (2004).
тривать как предшественников современных. По неустановленным причинам меловые ТЦС бесследно исчезли.
В кайнозое ТЦС сформировались на основе злаков, причём независимо, как минимум в трёх местах.
Первые по времени, в эоцене, ТЦС сформировались в Южной Америке, их эдификаторами были неполнозубые и вымершие отряды южноамериканских копытных — ксенунгуляты (Xenungulata) и нотоунгуляты (Notoungulata), а позднее — и крупные грызуны. В Северной Америке ТЦС начинают формироваться в олигоцене, а в миоцене этот процесс захватывает и Евразию с Африкой. Эдификаторами этих ТЦС являлись и являются парно- и непарнокопытные и хоботные. И, наконец, в неогене ТЦС начали формироваться в Австралии, их эдификаторами были травоядные сумчатые и крупные страусы, истреблённые человеком. Возможно, ТЦС формировались и в других местах: в Новой Зеландии (эдификаторы — страусы моа), на Альдабре (эдификаторы — черепахи).Процесс формирования травяных биомов и эволюции таксонов-эдификаторов шёл параллельно. Так, хоботные дали вспышки таксономического разнообразия в начале олигоцена, в миоцене, в первой половине плиоцена (рис. 4.41) — когда травяные биомы начали формироваться; после их становления разнообразие хоботных снизилось. Хоботные не просто осваивали новую адаптивную зону, они её формировали.
О ведущей роли крупных травоядных в возникновении ТЦС свидетельствует также то, что после их уничтожения исчезают и ТЦС. Весьма показательно также и то, что голоценовое вымирание крупных травоядных, являвшихся эдификаторами тундростепи, не сопровождалось столь же масштабными вымираниями в других блоках, за исключением непосредственно связанных с крупными травоядными. Большинство видов, входивших в ТЦС, существует и поныне, кое-где в Северной Америке и Сибири сохранились даже остатки этих ЦС в виде реликтовых островков холодных степей.
Еще по теме Примеры формирования ценотических систем:
- Ценотическая система Ценотическая система — что это такое?
- Ценотическая система в изменчивом окружении — меняется поток энергии через ценотическую систему
- Ценотическая система в стабильном окружении
- Эволюция ценотических систем
- Причины эндогенного развития ценотических систем
- Смена одной ценотической системы другой
- Пути преобразования ценотических систем
- Ценотические системы пастбищного и консортного типов
- Границы между сообществами и ценотическими системами
- Ценотическая система и географическая изменчивость ценофильных видов
- 3.3.3. формирование «систем» дифференцировочных условных рефлексов
- 3.6.5. Характеристика генотипа как сбалансированной по дозам системы взаимодействующих генов 3.6.5.1. Значение сохранения дозового баланса генов в генотипе для формирования нормального фенотипа
- Примеры адаптаций
- Эколого-ценотические стратегии вилов и популяций растений
- Примеры видообразования
- 6-5. Простые примеры применения диатропики
- 8-11* Пример связи эмбриологии и палеонтологии
- Примеры включения навоза в систему удобрения
- Третий пример (пределы точности измерения)
- 3.4. Примеры априорного подхода