Эколого-ценотические стратегии вилов и популяций растений
Понятие стратегии растений вытекает из общей теории адаптации живых организмов. В самом общем виде стратегию можно определить как интегральную, генетически обусловленную совокупность признаков и свойств растения, которые обеспечивают его популяциям способность существовать в определенных эко- лого-фитоценотических условиях (Миркин, Наумова, 1998).
Близкое по смыслу определение приводят И. Царика и др. (2001): стратегия - это совокупность приспособлений, свойств и особенностей, которые проявляются в процессе реализации генотипа особей в изменяющихся условиях среды обитания и обеспечивают их продолжительное существование, способность занимать свободные экологические ниши, переживать стресс и восстанавливать свою структуру и функции. Эти авторы считают также необходимым использовать понятие «популяционная тактика* как способность реагировать на конкретные биологические и экологические факторы. В сущности, как синоним к термину «стратегия* О.В. Смирнова (1987) предложила термин «поведение*, но последний используется в зоологии совсем в другом смысле, и поэтому это предложение не нашло поддержки у специалистов.Существует довольно много подходов к классификации реализуемых видами растений и популяциями вариантов стратегии.
При анализе логистического уравнения роста популяций была высказана идея, что отбор во флюктуирующей среде благоприятствует увеличению параметра г, а нахождение популяции в условиях стабильной природной среды - увеличению К. На этой основе Мак Артур и Уилсон ( McArthur, Willson, 1967) предложили концепцию г- Х-отбора, которая получила широкое распространение в геоботанической литературе. Она дозволяет дать достаточно четкую характеристику биолого-экологическим свойствам видов растений. В двух своих крайних вариантах виды растений по их стратегии могут характеризоваться следующим образом:
- г-виды - это некрупные растения, преимущественно однолетники или двулетники, у которых основная масса органических веществ направляется на формирование репродуктивных органов. Ориентация жизненных процессов на воспроизводство - их главная черта;
- К-виды - это крупные растения: деревья, кустарники и многолетние травы с медленным ростом. К генеративному размножению они приступают поздно и затрачивают на репродукцию сравнительно малую часть своей фитомассы. Главной чертой их биологии является стратегия выживания.
В соответствии со своими биологическими особенностями г-виды занимают нарушенные местообитания с частично пустыми экологическими нишами, а К-виды произрастают в кл и маисовых, равновесных сообществах. В критические эпохи существования биосферы, когда массово нарушаются климаксовые сообщества, именно с этим может быть связано вымирание крупных форм растений и животных. Е. Пианка (1970) показал, что между крайними г- и АГ-видами существует ряд переходных форм, что ведет к формированию континуума г ...К- стратегий. Реально ни один из видов растений не реализует какой-либо стратегии в чистом виде, имеется их определенное сочетание, и оно индивидуально для каждого вида растения (Домбровский, Грунтфест, 1994). Тип стратегии - характеристика относительная. «О каждом конкретном организме можно говорить как о r-стратеге или К-стра- теге только по сравнению с другими организма мл *, — подчеркивал Пианка (1981:138).
При своей определенной прогрессивности концепция Г-...К- стратегий вызвала в свой адрес ряд критических замечаний. Еще Деметриус (Demetrius, 1977) отметил, что эта концепция пренебрегает комплексностью онтогенетических явлений у растений и поэтому неадекватна их природе. К этому можно добавить и ряд других соображений. Фактически даже в равновесных популяциях иа фоне конкуренции генотипов в условиях AC-отбора может наблюдаться как увеличение, так и уменьшение параметра К. Крайние г- иК-типы, как, продемонстрировал Пианка(1981), могут быть неэффективными для адаптации видов к эколого-фито- ценотическим условиям, и тогда начинают отсекаться стабилизирующим отбором. На таком фоне у видов растений формируется четкая тенденция к сочетанию признаков г- и 1С-стратегий.
Не всегда плодовитость видов, положенная в основу концепции г-, К-стратегий, является решающей для выживания и эволюции. Более важной может оказаться сохраняемость семян в почве и выживаемость всходов. Концепция г-,..#-стратегий игнорирует тактики защиты растений, которая может иметь решающее значение для выживаемости видов. Эту концепцию трудно использовать при анализе материала широкой географической представленности. Известно, что растения теплого климата характеризуются общей тенденцией к формированию более крупных жизненных форм и к продуцированию крупных семян. Следует иметь в виду, что когда виды растений попадают в новые условия произрастания, они могут резко изменять тип своей стратегии. Репродуктивное усилие у однолетних многолетних растений может в таких случаях изменяться в амплитуде от 0 до 50%.В связи с очевидными теоретическими слабостями концепции r-...K-отбора не удивительно появление работ, авторы которых стремились сформулировать теории, более адекватные сущности биологических явлений. Широко известны системы Л.Г. Раменского (1938) и Грайма (Grime, 1979), Р. Уиттекера (1979, 1980), Дюринга (During et al., 1985), Б.М. Миркина (1986) и ряда других авторов (табл. 10.1). Разработка моделей стратегии видов стала достаточно модным делом. Поскольку адаптационные синдромы исключительно многообразны, то стратегий, по существу, столько, сколько видов растений. Но некоторые из них по своему содержанию близки друг к другу, отличаясь лишь небольшими деталями, что позволяет объединять стратегии в некоторое количество основных базовых типов.
По О.В. Смирновой (1987), интегральными базовыми свойствами растений являются три:
- конкурентоспособность, проявляющаяся в способности подавлять другие виды за счет высокой жизнедеятельности;
- фитоценотическая толерантность, представляющая способ ность длительно существовать на территории, которая контролируется конкурентоспособными видами;
- реактивность, состоящая в способности быстро захватывать освобождающиеся свободные территории.
Таблица ЮЛ.
Подходы разных авторов к установлению экологоцено- тических стратегий растенийАвтор
Выделяемые стратегии
Мак Артур и Уилсон (McArthur, Willson, 1967)
г-виды, ориентированные на репродукцию, /(-виды, ориентированные на выживание
В. Пианка (1970)
Л.Г. Раменский (1938)
Дж. Грайма (Grime, 1979)
Р. Уиттекер (1980)
Кэдмон и Шмида (Kadmon, Schmida, 1990)
Б.М. Миркин (1986)
A...K-континуум
Три базовых стратегии; виоленты, патиенты и эксплеренты
Повторное открытие стратегий, установленных Л.Г. Раменским под другими названиями: Я-виды (рудеральная стратегия = эксплеренты), С-виды (конкурентная стратегия = виоленты), S-виды (стратегия стресс-устойчивости = патиенты)
/(-стратегия соответствует максимальной плотности насыщения популяции особями, (.-стратегия отвечает нижней плотности насыщения и г-стратегия является флюктуирующей между этими полюсами
Пять типов демографических стратегий, отличающихся плотностью популяций, способностью формировать почвенный банк семян и типом распространения семян
Пять типов стратегии: /(-виоленты, S-экотопические патиенты, 5*-фитоценотические патиенты, Я-эксллеренты, Я*- фитоценотические эксплеренты
В соответствии с наличием этих свойств тот или иной вид относят, по О.В. Смирновой, к одной из перечисленных выше категорий. Из всех попыток развития исходной идеи о стратегиях видов эта оказалась наименее удачной, поскольку положенные в ее основу критерии относительны и неопределенны. Это в первую очередь касается конкурентоспособности и толерантности видов. Так, например, Aegopodium podagraria — действительно, толерантный вид по отношению к ряду широколиственных древесных пород, но она же - ярко выраженный конкурентный вид по отношению к ряду напочвенных зеленых мхов и даже по отношению к весенним эфемероидам, которых она быстро вытесняет их из своих популяционных полей.
Типы выделяемых стратегий, естественно, оказываются различными в зависимости от того, какие свойства видов и популяций кладутся в основу системы и на каких принципах выполнена
редукция многопризнаковости популяций.
В целом, разработка концепции жизненных стратегий видов растений шла от одномерного к многомерному подходу (Whittaker, Goodman, 1979, Болюх, 1996). При этом многими авторами было показано, что «чистых стратегий* при любом их определении очень мало и, как правило, виды растений реализуют промежуточные стратегии (Демидова, Уланова, 2006, Ильина, 2006 и др.).В настоящее время ведется активная работа по созданию единого перечня признаков растений для оценки стратегий их жизни (Weiher et al., 1999). Количество таких признаков колеблется от 50 до 6-10 (McIntyre et al., 1999). Такие признаки, очевидно, должны быть связаны в определенные комплексы — синдромы признаков. По Бредшо (Bredshaw, 1969), оценка стратегии жизни видов растений должна включать в себя три главных показателя:
- характеристику вида с точки зрения его возможностей в отношении факторов экологической среды, то есть экологическую амплитуду вида в ее чистом виде;
- характеристику вида с точки зрения его взаимоотношений с другими видами;
- оценку приспособления вида к сообитанию с другими видами.
Развитие концепции стратегий жизни растений привело к важному открытию: отличаются не только стратегии разных видов растений, отличаются стратегии, которые реализуют разные популяции одного и того же таксономического вида растения. Один и тот же вид растения в разных условиях, представленный конкретными популяциями, может реализовывать разные стратегии жизни. Межпопуляционная подвижность стратегий реал но существует и проявляется, когда разные популяции одного вида растения оказываются в заметно отличающихся эколого-ценоти-
ческих условиях.
К. А. Куркин (2002) привел ряд примеров, как на лугах типичные эксплеренты фактически являются латиентами. С.А. Дубров ная (1998) показала, что земляника лесная Fragaria vesca в со сновых лесах реализует тактику патиента, на антропогенных местообитаниях - эксплерента. М.Р. Турбина и А.К. Махнев (1999) установили, что в условиях стресса популяции скерды кровельной Crepis teciorum переходят от r-стратегии к Я-стратегии.
М.В. Марков (1990) даже описывал случаи перехода одного и того же вида растений в ряду жизненных форм «однолетники — двулетники — многолетники* в зависимости от условий произрастания.В настоящее время отсутствует теоретически обоснованная комплексная методика определения того, какую именно стратегию в действительности реализует та или иная популяция. Тем не менее основные признаки, которые могут быть положены в основу оценки стратегий популяций уже достаточно понятны, если использовать базовую идею континуума r-...A’-стратегий об альтернативном вкладе органических веществ либо в размножение, либо в формирование вегетативных органов. Это позволяет распределять виды растений и их конкретные популяции в двухмерном пространстве на основании этих двух основных признаков, которые характеризуют стратегию растения (рис. 10.2).
На оси абсцисс откладывается размер фитомассы растений, на оси ординат - репродуктивное усилие. Это удобно при сравнении популяций одного вида или популяций видов растений, ве-
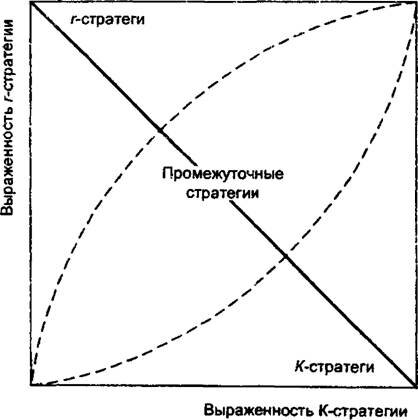
Рис. 10.2. Пространство распределения популяций в континууме двух признаков: выраженность/Г-стратегии (фитомасса особи) и г-стра-
тегии (репродуктивное усилие)
личина фитомассы особей которых отличается друг от друга только на один-два порядка. В случае сопоставления очень разноразмерных растений (например, деревья и однолетние травы) целесообразнее использовать логарифмическую шкалу или, что проще, принимать максимальный вес особи в выборке популяций за 100%, а остальные веса представлять в процентах от этой величины.
На оси ординат откладываются значения репродуктивного усилия. Оно варьирует от 0 до 100%, и прибегать к преобразованию шкалы в этом случае нет надобности. Главная диагональ квадрата континуума г-.. ./С-стратегий с помощью эллипса делится на три равные части. Зона эллипса отвечает пространству растений промежуточных стратегий, левая верхняя зона - растениям г-стратегий, правая нижняя зона - растениям /Г-стратегий. Такой метод позволяет достаточно объективно, на основе количественных признаков сопоставлять как стратегии видов растений, так и их отдельных популяций.
Так, Ю.А. Злобин и В.М. Кохановский (1991) при исследовании десяти популяций лука медвежьего Allium ursinum из разных местообитаний обнаружили (рис. 10.3), что на природных охра няемых территориях популяции (популяции 1, 2, 7) реализуют К-стратегию, но по мере нарушения фитоценозов, наблюдающе- госявзонах рекреации и зеленой зонег. Сумы у популяций (3,10,
- усиливается тенденция к r-стратегии. Популяции Allium ursinum в пространстве диаграммы явно образовывали отчетливый континуум стратегий. На его протяжении средняя фитомасса генеративных особей уменьшалась примерно в 3 раза, а репродуктивное усилие возрастало в 2 раза. Способность популяций гибко менять стратегии является адаптивным механизмом, расширяющим их экологическую нишу. Имеются аналогичные данные и по другим видам растений.
Сходный, но чисто эмпирический подход позже был разработан Вестоби (Westoby, 1993), который предложил ординировать популяции по трем осям (треугольник LHS), отображающим основные морфометрические признаки растений (площадь листьев, фитомассу, высоту, вес семян и др.).
Информативной и наглядной оказывается комплексная оценка своеобразия каждой отдельной популяции сразу по широкому набору признаков особей популяции и признаков самой популяции. Это удобно осуществлять методом лепестковых (круговых)

Рис. 10.3. Положение десяти популяций лука медвежьего Allium ursinum на фрагменте диаграммы -стратегий. По мере ухудшения условий произрастания происходит смена типа стратегии популяций в сторону усиления у них г-стратегии диаграмм
На рис. 10.4 представлены такие диаграммы для четырех популяций птицемлечника зонтичного Ornithogalum umbellatum, которые показывают, что эти четыре популяции неидентичны как по морфологическим параметрам отдельных особей, так и по структурным популяционным показателям (Кричфалуший, Мезев-Кричфалуший, 1994). Тот факт, что у одного и того же вида растения разные популяции могут осуществлять разные стратегии жизни, адаптируясь к конкретным условиям произрастания, является важным открытием в популяционной экологии растений.
Безусловно, существует и дифференциация стратегий между особями одной и той же популяции. Е.Л. Любарским (1985) описаны так называемые функциональные группы растений в фито популяции:
- растения, выполняющие функцию прироста фитомассы;
- резервная группа, обеспечивающая устойчивость популяции при неблагоприятных условиях;
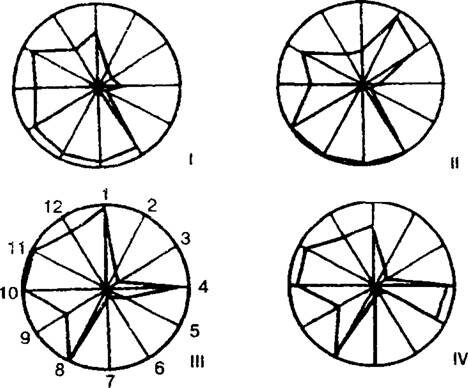
Рис. 10.4. Разные варианты эколого- ценот и ческой стратегии
птицемлечника зонтичного:
I--IV - четыре самостоятельных популяции; 1 - общая фитомасса, г/мг; 2 - плотность популяции, шт./0,25 м2; 3-фито- масса генеративных растений, г/мг; 4 - количество семян, шт./особь; 5 - ко-личество семян, шт./мг; 6 - количество луковичек, шт./особь; 7 - количество луковичек, шт./м2; 8 - долевая площадь листьев, см2/г; 9 - отношение фитомассы надземных органов к общей фитомассе, г/г; 10 - отношение фитомассы подземных и надземных органов, г/г; 11 - отосинтетическое усилие, г/г; 12-репродуктивное усилие, % (поВ.Е. КричфалушийиГ.М. Мезев-Кркчфалуший, 1994)
- группа, реализующая функцию вегетативного или генеративного размножения.
Таким образом, варьирование стратегий жизни - это не только свойство видов растений, но свойство их популяций и даже групп особей.
Еще по теме Эколого-ценотические стратегии вилов и популяций растений:
- Популяции как форма существования вилов живых организмов
- НЕКОТОРЫЕ ЭКОЛОГО-ФИЗИОЛОГИЧЕСКИЕ ПОКАЗАТЕЛИ, ХАРАКТЕРИЗУЮЩИЕ ВОДНЫЙ РЕЖИМ РАСТЕНИЙ
- Ценотическая система в изменчивом окружении — меняется поток энергии через ценотическую систему
- Ценотическая система Ценотическая система — что это такое?
- Признаки популяций растений
- Популяции культурных растений
- Биоразнообразие особей растений в популяциях
- Сущность и формы линамики популяций растений
- Разнообразие динамических процессов в популяциях растений разных жизненных форм
- Самоподдержание и популяциях растений
- Формы взаимоотношений между популяциями разных видов растений