Ценотическая система и географическая изменчивость ценофильных видов
Может показаться маловероятным, что при отсутствии барьеров ЦС способны сохранить свою целостность. Рассмотрим эту проблему на примере анализа географического распространения близких таксонов (внутривидовых форм или близких видов).
Итак, в ЦС ценофильный вид находится под мощным прессом стабилизирующего отбора в соответствии со своей нишей. Один и тот же вид может входить в состав нескольких ЦС, более того, он обычно входит. Но в другой ЦС вид имеет другую нишу (поскольку сообщества, в которые он входит, другие). Это объясняет широко известный феномен географической изменчивости и то, что часто географическая изменчивость носит не плавный (клинальный) характер, а вид можно разбить на некоторое число подвидовых таксонов с узкой зоной интерградации. И даже клинальный, а не дискретный характер изменчивости может быть артефактом из-за того, что при отборе и анализе изменчивости изначально предполагают клинальный её характер, границы же ЦС во внимание не принимают, и клина получается как результат формальной интерполяции. Часто один вид сменяет в пространстве близкий к нему (викарирующий). Обычно дискретность того или иного таксономического ранга интерпретируют как результат следующих событий: — превращение единого ареала вида в несколько изолированных участков (причиной этого в средних широтах полагают оледенение);
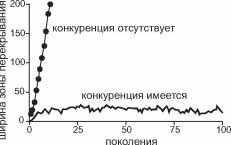
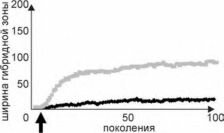
Рис. 4.33. Динамика ширины зоны Рис- 4-34. Динамика ширины гибридной зоны при
перекрывания ареалов в отсутствии беспрепятственной гибридизации и конкурентном
гибридизации. исключении в зависимости от радиуса расселения.
Стрелкой отмечен момент контакта, серая линия — будь- ший радиус, чёрная — меньший.
- аллопатрическое возникновение в изолятах в той или иной степени обособленных таксонов (вид, подвид);
- восстановление непрерывности ареала после устранения причины фрагментации;
- гибридизация (в случае подвидов) или конкурентное исключение (в случае видов) в зоне контакта.
Ширину зоны гибридизации рассматривают как функцию незавершённости процессов видообразования, её отсутствие (конкурентное исключение) — завершённым видообразованием.
Такую последовательность событий предполагают из-за широко распространённого представления о том, что при свободном генетическом обмене в зоне контакта должен установиться плавный градиент характеристик вида. Это представление кажется настолько очевидным, что его обоснование полагают излишним. Проанализируем это представление на простейшей модели (Грабовский, 1995). В модели можно устанавливать размер поля, радиус расселения, радиус поиска полового партнёра, плодовитость и влияние на спаривание «процента крови» одного или другого таксона, жизнеспособность гибридов. Модель можно запустить в двух вариантах: (1) один вид полностью конкурентно исключает другой (в каждой клетке поля может быть только один вид) и (2) виды не конкурируют друг с другом (в каждой клетке поля может быть как один вид, так и оба сразу).
При отсутствии спаривания в случае полного конкурентного исключения между видами устанавливается узкая зона перекрывания (ширина положительно зависит от радиуса расселения). В отсутствие конкуренции ширина зоны перекрывания линейно увеличивается, пока каждый вид не займёт всё поле (рис. 4.33). И тот, и другой результаты ожидаемы.
Если особи разных таксонов свободно спариваются, и их потомство полностью жизнеспособно, то в случае конкурентного исключения между ними устанавливается узкая зона перекрывания, ширина которой зависит от радиуса расселения (рис. 4.34). Даже в случае отсутствия конкурентного исключения между таксонами устанавливается узкая зона гибридизации, ни о каком плавном градиенте не может идти и речи.
Снижение жизнеспособности гибридов приводит к сужению зоны перекрывания, т. е. посткопуляционные механизмы эффективно препятствуют генетическому обмену. С уменьшением случайности спаривания (особи вида предпочтительно выбирают особей того же вида) ширина зоны перекрывания ареалов, наоборот, возрастает, это приводит к увеличению генетического обмена, если предпочтение в спаривании не абсолютно. Выше были рассмотрены случаи, в которых эффективные прекопуляционные механизмы отсутствовали. Эффективные же посткопуляционные механизмы имеются у всех ценофильных видов — это мощный пресс стабилизирующего отбора: гибридам места в ЦС нет. Это объясняет, как при отсутствии механизмов, предотвращающих гибридизацию, один и тот же вид может занимать разные экологические ниши в разных ЦС, как может вообще сфор-

Рис. 4.35. Формирование зоны перекрывания при свободном скрещивании, 100% смертности гибридов и низкой плодовитости через 200 поколений.
мироваться новая ЦС в отсутствии изоляции.
При разрушении ЦС (например, в антропогенных биотопах) посткопуляционные механизмы не действуют, что увеличивает зону перекрывания.
Рассмотрим также случай отсутствия конкуренции и прекопуляционных механизмов (свободного скрещивания), но низкой жизнеспособности гибридов. При низкой плодовитости между таксонами устанавливается узкая зона, ширина которой пропорциональна радиусу поиска полового партнёра. Появление этой зоны, по-видимому, связано с высокой смертностью гибридов. С увеличением плодовитости и/или радиуса поиска полового партнёра зона перекрывания быстро расширяется, и при достаточно высокой плодовитости даже при 100% смертности гибридов ареалы видов быстро полностью перекрываются (при тех же параметрах, что и на рис. 4.35, но плодовитости не 4, а 100 — за 12 поколений).
Перейдём к примерам из природы. Ограничимся наиболее широко известными, хрестоматийными случаями: кольцевой ареал большой синицы Pams major (рис. 4.36) и серой Corvus comix и чёрной С. согопе ворон (рис. 4.37). Оба этих случая рассматривают авторы практически всех учебников по теории эволюции как примеры аллопатрического видообразования и зон вторичного контакта.
Парадигма структурализма позволяет иначе трактовать эти случаи. Pams major major — европейский подвид, естественный ареал которого ограничен преимущественно буковыми лесами. Экспансия подвида на восток связана с освоением им антропогенных ландшафтов, в первую очередь населённых пунктов. Pams major minor — восточноазиатский подвид, экспансия которого на север также связана с освоением им тех же антропо-
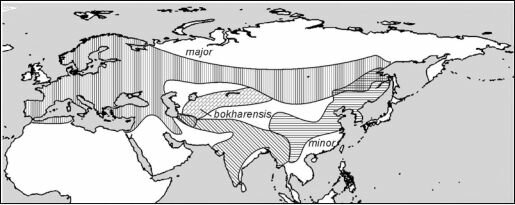
Рис. 4.36. Кольцевой ареал большой синицы. По Кейн (1968), из А.С. Северцова (2005, упрощено).
Надписаны названия подвидов. 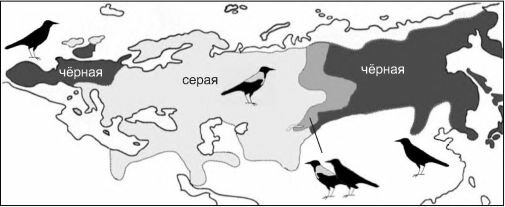
Рис. 4.37. Гнездовые ареалы и зоны гибридизации чёрной и серой ворон. По Блинову (1998), из
А.С. Северцов, 2005).
генных ландшафтов. Очевидно, что в обоих случаях при расширении естественного ареала подвида и вселении в антропогенные ландшафты и тот и другой виды вышли из-под контроля родных для них ЦС и вошли в состав формирующихся синантропних ЦС, сохраняющих ещё многие свойства группировок. Как чистым подвидам, так и их гибридам есть в них место, поэтому между подвидами в узкой полосе происходит гибридизация.
Серая ворона в естественных сообществах — вид свойственный преимущественно приречным районам, чёрная — вид, более свойственный лесам. Оба вида давно стали (каждый в пределах своего естественного ареала) одними из обычнейших синантропных видов.
Как и в случае с большой синицей, гибридизация происходит вне природных ЦС. Ширина зоны гибридизации в полном соответствии с моделью определяет подвижность особей: в Западной Европе виды более осёдлы, и зона гибридизации существенно уже.В обоих рассмотренных выше случаях таксоны аллопатричны — замещают друг друга в пространстве (викарируют). Рассмотрим теперь случай симпатричных видов. Всё представители семейства Fagaceae (буковые) являются ценофилами. Многие виды — эди- фикаторы различных ассоциаций, в том числе и климаксных, как в Северном, так и Южном полушарии, как в Старом Свете, так и в Новом. В Северном полушарии широко распространён род дуб — Quercus. Дуб обыкновенный, или черешчатый Q. гоЪиг является эдификатором климакса Восточно-Европейской ботанико-географической провинции (север Восточной Европы, на восток почти до Урала). В Западно-Европейской ботанико-географической провинции (территория Западной Европы, а также на юге Восточной Европы и Кавказа до Ленкорани включительно) эдификатором климакса является скальный дуб Q. petraeci, отличающийся почти сидячими желудями (рис. 4.38). Эти два вида близки друг к другу, их ареалы в значительной степени перекрываются: у обоих видов они, например, включают Британские острова, так что в Шервудском лесу Робина Гуда были оба вида. Там, где их ареалы перекрываются, Q. гоЪиг становится эдификатором предшествующих климаксу сериальных ассоциаций. На всём протяжении их ареалов это — «хорошие» виды. Согласно палеонтологическим данным, оба вида существуют, как минимум, на протяжении последних 10 млн. лет (с миоцена), всё это время их ареалы перекрывались на площадях в сотни тысяч квадратных километров, и никаких сомнений в их самостоятельности не возникало. Однако выяснилось, что в искусственных насаждениях (т. е. вне природных ЦС, ценофилами которых они являются) эти два вида неограниченно скрещиваются, давая вполне плодовитое потомство (White, 1981). После обнаружения этого факта начали говорить о том, что это — «полувиды».
Смена названий ничего в действительности не объясняет, т. е. не объясняет того, почему эти виды сосуществовали миллионы лет на одной территории и сохранили свою самостоятельность. С позиций структурализма ситуация с этими двумя видами настолько классическая, ясная и понятная, что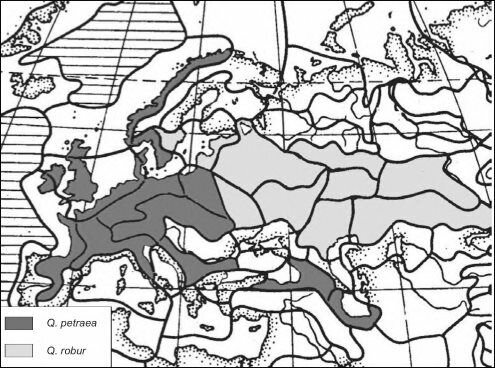
Рис. 4.38. ЦС, в которых эдификаторами климакса являются Quercus petraea и Q. robur. По
С.М. Разумовскому (1999).
так и просится в учебник: виды были раздельны в естественных сообществах, пока (и поскольку) были ценофилами. В искусственных посадках обычно ни о каких естественных ассоциациях говорить не приходится, это типичные группировки, поэтому оба вида дуба в них следует считать ценофобами, т. е. они выходят из-под контроля ценозов. Скрещивание, по-видимому, имеет место и в природе, но там нет ниши для гибридов, а есть лишь ниши Q. petraea и Q. robur, поэтому гибридные особи не выживают. В группировках же чётко очерченных ниш нет, что позволяет существовать как Q. petraea и Q. robur, так и их гибридам. Этот пример также показывает, что «хорошие» виды (а это — всегда ценофилы) могут существовать симпатрично, не обладая репродуктивной изоляцией. Их самостоятельность поддерживает их ценофильность.
Аналогичная ситуация описана и для австралийских Eucalyptus, для многи из которых известны межвидовые гибриды, которые «как правило отсутствуют в сиапыз насаждениях. Чаще они наблюдаются на площадях, изменённых деятельностью человека» (Вальтер, 1975: 179).
Потомство многих видов в искусственных условиях (т. е. вне ЦС) вполне жизнеспособно, что показывает отсутствие у них действенной репродуктивной изоляции на уровне видов. Например, гибриды соболя (Maries zibellina) и куницы (Maries martes) — киду- сы — вполне плодовиты, что многократно проверено в пушном разведении. Ареалы соболя и куницы широко перекрываются (куница доходит на восток до Оби, а соболь на запад до Печоры), но самостоятельность их несомненна. Примеров видов, свободно скрещивающихся в неволе (т. е. вне ЦС), но длительно сосуществующих в природе (т. е. в ЦС), очень много: все утки (Anatidae) и полорогие Bovidae, многие олени способны к свободному скрещиванию, их потомство вполне жизнеспособно, в случае уток это даже создаёт определённую проблему в зоопарках. Дополнительные примеры по позвоночным можно найти у С.С. Шварца (1980) и Е.Н. Панова (1989).
Концепция ценофильных и ценофобных видов позволяет понять существование морфологически отдельных групп особей (видов), для которых неизвестно половое размножение, т. е. основной критерий вида — общность генофонда — не применим. Вид, таким образом, не генетическое, а экологическое понятие.
Еще по теме Ценотическая система и географическая изменчивость ценофильных видов:
- Ценотическая система в изменчивом окружении — меняется поток энергии через ценотическую систему
- Ценотическая система Ценотическая система — что это такое?
- Географическая изменчивость ФПР
- Ценотическая система в стабильном окружении
- Примеры формирования ценотических систем
- Эволюция ценотических систем
- Причины эндогенного развития ценотических систем
- Ценотические системы пастбищного и консортного типов
- Пути преобразования ценотических систем
- Границы между сообществами и ценотическими системами