Ценотическая система Ценотическая система — что это такое?
Итак, многочисленные примеры показывают, что в некоторых случаях быстро (иногда практически мгновенно) образуются эндемичные виды и экосистемы. Поскольку образование новых видов может происходить весьма быстро, кажется вероятным, что развитие взаимодействий между видами может происходить по крайней мере столь же стремительно, а, может быть, и ещё быстрее.
Закономерности этого процесса, т. е. эволюция, возникновение и развитие экосистем — основное содержание концепции структурализма. Парадигма структурализма сравнительно малоизвестна, её изложения нет ни в одном учебнике или монографии, как в отечественных (если не считать фрагментарные её изложения в трудах континуалистов), так и, насколько мне известно, в иностранных. В то же время, с моей точки зрения, она адекватнее описывает структуру биосферы (и бентали, как её составной части). Поэтому, прежде чем рассмотреть структуру бентали, необходимо обсудить общие вопросы экологии. Желающие более углублённо ознакомиться с теорией структурализма могут обратиться к сборникам трудов, изданных издательством КМК (Разумовский, 1999, 2007; Жерихин, 2003).Размеры экосистем, возникающих в рассмотренных выше примерах, гораздо больше размеров отдельных биоценозов и сообществ. Фактически многие из таких экосистем очевидно образованы целым комплексом сообществ. Поэтому для таких крупномасштабных экосистем С.М. Разумовским был использован термин ценотическая система[††††††††] (Разумовский, 1969; Киселева, Разумовский Расницын, 1969; Киселева, Разумовский, 1971 и др.), в дальнейшем — ЦС. Новая ЦС может сформироваться лишь там, где возникает пространство, удовлетворяющее трём условиям:
— На данной площади не может существовать никакая из имеющихся ЦС, могущих попасть на неё.
- Размер площади должен быть достаточно велик для существования ЦС, а условия на ней предсказуемы.
- Оба предыдущих условия должны соблюдаться на протяжении времени, достаточного для формирования ЦС.
Относительно последнего условия — времени — можно отметить, что время, необходимое для формирования ЦС, возможно, уменьшается с: (1) увеличением отличий условий, в которых формируется ЦС, от условий системы-донора и (2) уменьшением числа видов, участвующих в её образовании.
Эти условия кажутся неконкретными и слишком строгими, а соответствующие им площади — редко встречающимися в природе. В действительности это не так — подобные площади возникают постоянно. Во всех таких случаях мы наблюдаем возникновение ЦС и обычно сопровождающий их очаг эндемизма (видов, ареалы которых ограничены только данным районом). Очевидно, что примерами подобных площадей являются:
- ультраабиссальные желоба посреди абиссальных равнин;
- изолированные глубоководные котловины (Северный Ледовитый океан, Японское и Средиземное моря);
- некоторые крупные и не очень крупные озёра (Байкал, Ньяса и т. п.), а также острова;
- достаточно крупные горы или горные системы посреди равнин.
Сложнее заметить, что этим условиям полностью удовлетворяют:
- высокие широты, после произошедшего в олигоцене - миоцене увеличения температурного градиента между экватором и полюсом (относится в равной степени к территориям и акваториям): в конце палеоцена поверхностная температура воды в Антарктике достигала 18°С.
- приэкваториальные районы, после произошедшего в то же время повышения температуры выше экологической амплитуды существовавших в этих местах ранее субтропических ЦС.
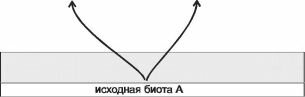
Рис. 4.26. Возникновение видов и биот с позиции структурализма.
Обратите внимание на то, что биоты формируются на отрезках плавного градиента. В отличие от рис. 4.10, на схеме полностью отсутствуют каких-либо преграды между формирующимися биотами. Нет даже областей увеличенных градиентов. Границы между биотами тем не менее резкие. Дополнительные пояснения в тексте.
| N | |
| биота А | биота Б |
градиент среды
| биота А | биота Б | биота В |
Ни в одном из приведённых выше примеров (а число их можно значительно увеличить) не возникала преграда, ареал формирования новой ЦС всегда оставался связанным с ареалом материнских ЦС.
В последних двух случаях формирующиеся ЦС имели общие границы с ЦС-донорами на протяжении тысяч километров! Поэтому рис. 4.10 не то чтобы не верен, но подчёркивает второстепенную, совершенно не обязательную причину видообразования — фрагментацию ранее единой биоты.При этом действительно важная причина, имеющая гораздо большее значение и распространение, — возникновение на обширных площадях условий, выходящих за пределы, в которых существующие ЦС способны нормально функционировать, — на нём никак не отражена. Поэтому, возможно, более правильной следует считать совершенно другую схему возникновения биот (видов) — изображённую на рис. 4.26.
Итак, ЦС возникают на достаточно однородных площадях, условия на которых выходят за пределы экологических амплитуд ЦС, которые могут на эту площадь попасть. Существование какого-либо барьера при этом вовсе необязательно, по-видимому, его обычно и нет — ареал формирующейся ЦС остаётся широко сообщающимся с ареалами старых ЦС. Но, поскольку условия в ареале формирующейся ЦС существенно отличаются от условий в ареалах старых, между их ареалами существует градиент условий. Граница между ЦС устанавливается в области таких значений внешних факторов, при которых конкурентоспособность (способность занять и удерживать территорию) контактирующих ЦС равна.
Рассмотрим в эволюционном масштабе времени процесс возникновения ЦС на ранее безжизненной площади. Еще раз можно напомнить случай Арала, показывающий, что «эволюционный масштаб времени» может быть всего лишь парой десятилетий. Для рассмотрения действительно общих экологических закономерностей не важно, акватория это или территория. Рассмотрим крайний случай: виды, заселяющие рассматриваемую площадь не имеют никаких адаптаций ни друг к другу, ни к специфическим условиям новой площади. Очевидно, что при первом же взаимодействии видов выявятся их макрониши (одни — автотрофы, другие — консументы первого порядка, третьи — хищники и т. п.). В пределах макрониш одни виды станут более обильными и обычными просто случайно, либо из-за того, что они окажутся более конкурентоспособными, нежели другие.
Такие виды будут в той или иной степени формировать окружающую среду, снижая её изменчивость (хотя бы лишь фактом своего наличия). Это приведет к возникновению ассоциаций видов. Чем более сильным эдификатором окажется вид (или группа видов), тем больше такая ассоциация будет приближаться к консорции. При достаточно высоком разнообразии условий на рассматриваемой площади в других её частях будут другие условия, в которых доступные ресурсы будут более подходящими для других видов. Они тоже будут формировать свои ассоциации. Например, одни ассоциации будут возникать на рыхлых грунтах, другие — на скалах. Одни на переувлажнённых почвах, другие — на песке или сухих известняках. Т. е. первоначально бесструктурная в биотическом отношении площадь будет становиться всё более структурированной по мере формирования ассоциаций.В силу чисто случайных причин (булыпая распространённость на данной площади одних условий и меньшая — других) одни из ассоциаций окажутся распространённее
«По мере образования биоценозов с устойчивой структурой, растения более чётко распределяются по различным местообитаниям» (Толмачев, 1974: 212).
других. Они, с одной стороны, окажут более сильное влияние на окружающие, просто в силу того, что они более распространены, т. е. являются предсказуемым элементом окружения. С другой стороны, наиболее распространённые ассоциации имеют больше возможностей для более тонкой и совершенной коадаптации входящих в него видов. Такие ассоциации оказывают всё более сильное влияние на остальные как всё более предсказуемый компонент среды. Процесс происходит, таким образом, с положительной обратной связью. В результате интеграция ЦС быстро растёт.
Ассоциации формирующейся ЦС, весьма вероятно, имеют несбалансированный обмен, поскольку для тонкой подгонки биологии отдельных видов просто не было времени. Это изменяет место, где находится сообщество (его биотоп): происходит накопление продуктов жизнедеятельности и/или перепотребление имевшихся изначально в данном месте ресурсов и т.
д. Мидиевые банки заиляются из-за отложения фекалий и псевдофекалий. Озера мелеют из-за отложения ила и торфа. На бесплодных песках откладывается гумус. В результате ассоциация изменяет свой биотоп таким образом, что в этом месте более конкурентоспособным оказывается другая ассоциация, которая её сменяет. Эту смену ассоциаций под действием причин внутренних, появившихся в результате их функционирования, называют экогенетической сукцессией. Движущей силой экогенетической сукцессии, как и любой другой сукцессии, является не- скомпенсированность воздействия популяций друг на друга. Необходимо отметить, что такие изменённые биотой биотопы могут существенно отличаться от имевшихся изначально на данной территории биотопов, т. е. функционирование биоты приводит к росту разнообразия условий в данной местности. Это обеспечивает возможность роста биоразнообразия и тем булыпе, чем булыпе ассоциаций формируется.дуб с орешником и волосистой осокой ?
ель с майником двулистным ?
политрихум обыкновенный ?
сфагнум Гиргензона ?
вейник седеющий ?
сабельник болотный
Рис. 4.27. Сукцессия зарастания водоёма в Московском ботанико-географическом районе. Ассоциации видов названы по доминанту. По
С.М. Разумовскому (1981).
Каждая ассоциация меняет свой биотоп одним единственным определённым образом (миди- евая банка заиляет грунт, а не откладывает гумус и не способствует зарастанию озера). Новому состоянию биотопа соответствует другая ассоциация.
Эту последовательность сменяющих друг друга во времени ассоциаций называют сукцессионный ряд (sere), а ассоциациями, входящие в этот ряд, — сериальными ассоциациями, или сообществами. Каждому достаточно отличному и распространённому исходному состоянию абиоты соответствует своя пионерная ассоциация и идущий от неё сукцессионный ряд. Сериальные ассоциации, подобно стадиям онтогенеза, неустойчивы во времени.
Примером сукцессионного ряда являетсяи зарастание водоёмов (стариц, низовых болот, мелких речушек и т.
п.) на территории Московского геоботанического района (рис. 4.27).Ассоциации, входящие в ЦС, образуются не сразу и не одновременно. Анализ структуры изученных ЦС и их сукцессий показывает, что ранние стадии сукцессий часто образованы более древними эволюционно таксонами или же более древними ассоциациями. Так, в гидросериях (сукцессионных рядах наземных ЦС, образованных на переувлажнён- ных субстратах) первые или одни из первых стадий образуют мхи. Ранние стадии восстановления современных рифов герматипных кораллов образованы водорослями — бурыми и красными известковыми. Развитие миоценовых коралловых рифов северо-восточной Атлантики, описанное в этой главе ниже, начинается с красных известковых водорослей, а как отмечено в главе 3, всегда (в палеозое, мезозое и кайнозое) при формировании сообществ мелководных рифов кишечнополостных геологически им предшествовали рифы красных известковых водорослей. Возможно, мы имеем дело с неким аналогом известного принципа «онтогенез повторяет филогенез».
Сукцессионные ряды формируются только для достаточно распространённых стартовых условий абиоты. Редко встречающиеся сочетания факторов не имеют своих сукцессионных рядов и такие местообитания осваивают ассоциации, сформировавшихся в биотопах со сходными условиями. Если в ареале ЦС возникает достаточно обширная площадь с условиями ранее не встречавшимися или редкими, то вполне может возникнуть новый сукцессионный ряд или модифицироваться существующий. Иными словами, несмотря на существование жёсткой сукцессионной последовательности, ЦС не следует понимать как окостеневшую структуру, она способна меняться вслед за изменением условий её существования.
Выше рассмотрена идеализированная ситуация, в которой ЦС формируется из видов, не имеющих никаких адаптаций ни друг к другу, ни к специфическим условиям новой площади в отсутствии преемственности с ЦС, существовавшей в этом месте ранее. В действительности такая ситуация невероятна. В формирующейся биоте обычно существуют виды, преадаптированные к биотопам новой площади, они выступают «ядрами конденсации» образующихся сообществ. Ещё эффективнее роль таких ядер будут выполнять группы коадаптированных видов. Весьма вероятно, что эффективность роли ядер растёт нелинейно с увеличением числа входящих в них видов. Поскольку, обычно, одна ЦС образуется из другой, число таких ядер во вновь формирующихся ЦС должно быть велико.
Процесс формирования новой ЦС на основе уже существовавших (одной или нескольких) происходит так же, как и в случае отсутствия такой преемственности. Основное отличие — наличие большого числа уже сложившихся и коадаптированных комплексов видов. Часто в состав формирующейся ЦС могут даже входить в почти неизменном виде ассоциации материнских ЦС. Эти «ядра конденсации» должны существенно ускорять формирование новых ЦС. В принципе, новая ЦС может целиком состоять из «старых» видов, но обычно часть видов специфична для неё, впрочем обычно не для отдельной ЦС, а для группы близких, т. е. им присуща та или иная степень эндемизма.
В ходе сукцессии несбалансированность обмена ассоциаций уменьшается. Это связано с тем, что сменяющие друг друга в ходе сукцессии ассоциации занимают одно и тоже место в пространстве, а количество поступающей на это место энергии постоянно. Всякая несбалансированность — это потери. Ассоциация, меньше теряющая энергию, оказывается в выигрыше. Также в результате сукцессии биотоп конкретной ассоциации видоизменяется таким образом, что отличия его от других уменьшаются. В результате сукцессионные ряды, начавшиеся в разных условиях с разных пионерных ассоциаций, сливаются друг с другом, постепенно приходя к ассоциации, которая в данных абиотических условиях (климат, глубина, обеспеченность пищей, освещённость и т. д. и т. п.) практически сбалансирована. Такую ассоциацию называют климаксной ассоциацией, или просто климаксом. В климаксе воздействие каждой популяции компенсирует воздействие остальных популяций. Климаксная ассоциация находится в состоянии динамического равновесия со своим биотопом. В результате изменение биотопа, если и происходит, то несущественно с точки зрения самого климаксной ассоциации. Например, климаксовый коралловый риф может сколь угодно расти в размерах, но как был климаксом, так климаксом и останется. Единственный признак климакса — отсутствие у него внутренних причин для изменения. Фактически более правильно определить ЦС как совокупность сукцессионных рядов (сериальных ассоциаций и климакса).
В принципе нет ничего невероятного в том, что на территории одной сукцессионной системы сформируется сколько угодно климаксов. Весьма вероятно, что при формировании ЦС и начинает формироваться несколько климаксов. Несколько климаксов сосуществуют и при превращении субклимакса в климакс, и при удлинении сукцессионных рядов. Однако, поскольку каждый климакс и его ЦС занимают определённую площадь, между разными климаксами будет происходить конкурентная борьба за занимаемую площадь. Поэтому, если сосуществующие климаксы требуют примерно одинаковых условий, то рано или поздно останется один. Иная ситуация будет наблюдаться, если сосуществующие климаксы требуют совершенно различных условий. Например, климакс на рыхлых грунтах является климаксом в совершенно иных условиях, нежели климакс рифов. Такие климаксы могут сосуществовать неограниченно долго.
При нарушении ассоциации внешними факторами возможны два варианта. Если вместе с ней нарушается и её биотоп, то сукцессия начинается с ассоциации, соответствующей новому состоянию биотопа.
Если нарушение затрагивает только ассоциацию, оставляя её биотоп (в том виде, в который его модифицировали существовавшие в данном месте ассоциации) нетронутым, происходит демутационная сукцессия — сравнительно быстрая последовательная смена нескольких недолговечных ассоциаций. Каждой достаточно продвинутой коренной ассоциации экогенетической сукцессии может соответствовать один или несколько демутаци- онных сукцессионных рядов, состоящих из производных ассоциаций. Совокупность де- мутационных сукцессионных рядов и коренной ассоциации называют демутационный комплекс. Таким образом, экогенетическая сукцессия в действительности состоит из последовательности не ассоциаций, а демутационных комплексов. В качестве примера на рис. 4.28 приведены демутационные сукцессии климакса Московского ботанико-географического района. Обратите внимание на то, что имеются две начальные стадии сукцес-
дуб с орешником и волосистой осокой *
ель с волосистой осокой
ельник-кисличник

ель с пальчатой осокой
берёза с волосистой ожикой
ельник-черничник
берёза с вейником тростниковидным
луг душистого колоска
Рис. 4.28. Демутационные сукцессии климакса в Московском ботанико-географическом районе. По С.М. Разумовскому (1981).
зарастание гарей
зарастание вырубок
сионных рядов. Один сукцессионный ряд начинается в том случае, если почвенный покров не поврежден (после верховых пожаров, вырубок, ветровалов и т. п.). Другой сукцессионный ряд начинается после повреждения почвенного покрова (после низовых пожаров, хищнических рубок с использованием тяжёлой техники и трелёвки бревен, в процессе которых почва часто сдирается до материнской породы). После прохождения нескольких стадий эти два сукцессионных ряда сливаются в один.
Коренная ассоциация одинаково хорошо возобновляется как на собственном фоне, так и на фоне одного или нескольких производных ассоциаций. Ассоциации же демута- ционного ряда не способны возобновляться на собственном фоне: после естественного от возраста умирания их видов-эдификаторов одна ассоциация сменяет другую, а потому их существование ограничено продолжительностью жизни одного поколения растений древесного яруса. В демутационных ассоциациях с древесным ярусом отсутствует подрост соответствующих видов деревьев. Это явление достоверно показано для всех изученных в этом отношении демутационных ассоциаций (Разумовский, 1981, 1999, 2006).
Широко известен феномен тропических лесов, в значительной степени обуславливающий их высокую мозаичность: под деревьями обычно отсутствует подрост того же вида. Для его объяснения было предложено несколько теорий, но ни одна из них не объясняет удовлетворительно все факты. Вполне вероятно, что тут имеет место демутационная сукцессия.
Это различие одновременно позволяет естественным образом разделить биоценоз и его биотоп. Под биоценозом следует понимать не только саму биоту, но и сумму всех отклоняющих воздействий, оказываемых ею на абиоту и бесследно исчезающих после устранения биоты. Например, пониженная гидродинамика в глубине зарослей макрофитов является свойством сообщества, а не его биотопа, поскольку обусловлена исключительно действием талломов макрофитов.
В сообществах наземных растений сукцессии были детально рассмотрены С.М. Разумовским (1981). Он показал, что сукцессионные ряды различаются в зависимости от начальных условий, но все сукцессии приводят к климаксу, обычно одному в ЦС. Быстрее всего проходит демутационная сукцессия, но и её продолжительность не может быть меньше суммарной продолжительности жизни эдификаторов каждой стадии. Поскольку эдификаторы всех сериальных ассоциаций, за исключением нескольких пионерных, — деревья, очевидно, что минимальное время даже демутационной сукцессии порядка 1000 лет. Время прохождения экогенетических сукцессий очевидно гораздо больше (тысячелетия, часто много тысячелетий), поскольку при их прохождении происходит не только смена доминантов, но и видоизменение биотопа. Как пример приведена сукцессионная система Московского геоботанического района (рис. 4.29). Очевидно, что время, прошедшее после разрушения человеком сообществ тундростепи, во многих случаях явно недостаточно для завершения экогенетических сукцессий. Интересующиеся детальным разбором приведённой схемы и аналогичных схем других районов могут обратиться к уже цитированному «Полному собранию сочинений...» С.М. Разумовского (2007).
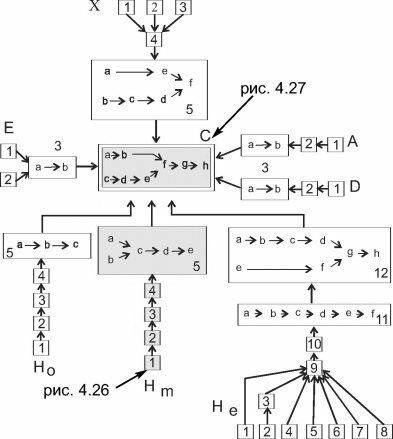
Рис. 4.29. Сукцессионная система Московского ботанико-географического района. По С.М. Разумовскому (1981)
Серым залиты части сукцессионной системы, рассмотренные на двух предыдущих схемах. С — климакс, сукцес- сионные ряды обозначены прописными латинскими буквами, стадии этих рядов — арабскими цифрами и прямоугольными рамками, а стадии демутационных рядов — строчными латинскими буквами. В случае двух демутаци- онных рядов в одном демутационном комплексе верхний из них — вырубки, ветровала, верхового пожара и других воздействий, уничтожающих верхний ярус сообщества, но не затрагивающих подстилку, а нижний — с нарушением подстилки, он часто возникает после низовых пожаров, но может быть вызван и любым другим воздействием, уничтожающим не только древостой, но и подстилку (трелевка с кронами, сбор подстилки населением, выпас в лесах, на крутых склонах и т. п.).
Еще по теме Ценотическая система Ценотическая система — что это такое?:
- Ценотическая система в изменчивом окружении — меняется поток энергии через ценотическую систему
- Ценотическая система в стабильном окружении
- Эволюция ценотических систем
- Примеры формирования ценотических систем
- Пути преобразования ценотических систем
- Причины эндогенного развития ценотических систем
- Смена одной ценотической системы другой
- Границы между сообществами и ценотическими системами
- Ценотические системы пастбищного и консортного типов
- Ценотическая система и географическая изменчивость ценофильных видов
- 11. Теплота. Что это такое?
- что ПРОИСХОДИТ ВНУТРИ КЛИМАТИЧЕСКОЙ СИСТЕМЫ
- Эколого-ценотические стратегии вилов и популяций растений
- 1-2. Что такое эволюция