12.3.2. Стрессоры и стресс
Неблагоприятный фактор, который вызывает у растения комплекс негативных процессов, снижающих его жизнеспособность и репродукцию, по предложению Г. Се лье (1992), называют стрессором, а само угнетенное состояние растения - стрессом.
Стресс у растений возникает в результате воздействия обширного комплекса эколого-ценотических неблагоприятных факторов, и его проявления также оказываются комплексными и разноуровневыми. Рецепторами восприятия стрессовых факторов, как первичные дискретные единицы живой материи, выступают отдельные особи растений. Вторичные проявления стресса изменяют популяционных параметры.
Обычно стрессовые воздействия не единичные и не разовые (хотя бывают и такие случаи). Это ведет к постепенному накоплению негативных последствий и формированию кумулятивного эффекта воздействия неблагоприятных факторов на популяции растений. За счет совокупности всех этих процессов популяции переходят в критическое состояние (Заугольнова и др., 1992), становятся малоустойчивыми и при дальнейшем нарастании или сохранении стрессовых условий выпадают из сообщества.
Следует различать стрессовое состояние особей растений и стрессовое состояние популяций.
На уровне особей под влиянием стрессоров происходят следующие изменения:
- снижается уровень продукционного процесса;
- уменьшаются размеры растения;
- замедляется или, напротив, ускоряется переход растений из одного онтогенетического состояния в другое;
- у генеративных особей однолетних растений снижается производство зачатков размножения, а в некоторых случаях оно прекращается вообще;
- у многолетних растений, находящихся в потенциально генеративном состоянии, в отдельные годы репродуктивный процесс отсутствует;
- можег изменяться жизненная форма растения.
На уровне популяций стрессоры сказываются следующим образом:
- за счет смертности особей, которые находятся в наиболее уязвимых онтогенетических состояниях, снижается численность растений в популяции;
- уменьшается популяционная плотность;
- онтогенетический спектр популяций становится неполночленным;
- снижается питал итет популяции за счет увеличения в ней доли угнетенных особей и снижения доли процветающих.
Согласно JI.A.
Животовскому (1998:126), «популяционный стресс - это значительное уменьшение приспособленности популяции под действием изменившихся факторов среды». Выживание популяций при стрессе может обеспечиваться двумя способами - сменой реализованной экологической ниши или удержанием экологической ниши (Усманов, 1991).Как общее правило, практически любой эколого-фитоценоти- ческий стресс ведет к подавлению продукционного процесса, и общий размер фитомассы растения снижается, В итоге происходит общая миниатюризация особей растений, при которой про порционально уменьшению размера общей фитомассы уменьшается размер всех частей растения. Но такое явление наблюдается нечасто. Его можно наблюдать, главным образом, в посевах в слу чае неоптимального загущения стояния культурных растений и создания популяции высокой плотности. Так, при анализе посева люцерны Medicago saliva с нормой высева 6, 8,12 и 14 кг семян на 1 га к фазе цветения по градиенту увеличения плотности посева наблюдалась общая миниатюризация растений (при исключении из рассмотрения краевых особей на опытных участках).
Общая структурная целостность особей растений в таких случаях остается высокой, приспособительное реагирование на стресс почти полностью отсутствует. Особи примерно все одинаково подвергаются миниатюризации.
Более типична и характерна в первую очередь для растений природных экосистем компенсаторная адаптация к стрессу, при которой наблюдается:
- уменьшение отдельных размерных показателей;
- стабильное сохранение их размера;
- увеличение отдельных размерных морфопараметров.
Такой внутренней перестройкой растения реагируют на стрессовую обстановку, а в некоторых случаях и приспосабливаются к ней. Число вариантов подобных трансформаций достаточно велико, так как зависит от жизненной формы растений и реализуемой им стратегии жизни. Биологическая интерпретация таких трансформаций непростая, поскольку у растений одновременно наблюдаются как адаптивная, так и стрессовая пластичность.
Первая повышает приспосабливаемость растения к неблагоприятным условиям, вторая отражает уровень ингибирования фи- зиолого-биохимических и формообразовательных процессов при стрессе.Ростовые и формообразовательные стрессовые изменения составляют единый комплекс, в котором имеют место и чисто стрессовая, и адаптационная компоненты. Общеизвестен «синдром избегания затенения* у растений. Он включает в себя изменение направления аллокации веществ, удлинение стебля, уменьшение разветвленности и изменение уровня репродукции. Как компенсирующая адаптация у таких растений одновременно существенно возрастает размер листовой поверхности, что, очевидно, обеспечивает выживание растений.
В целом, внешние воздействия на популяцию растений отличаются по характеру: одни нивелируют популяционную изменчивость и выравнивают особи между собой, другие - усиливают дифференциацию особей.
В стрессовой эколого-фитоценотической среде происходит сужение реализованных экологических ниш, и, чем дольше действует стрессовая обстановка на растения и их популяции, чем мощнее это воздействие, тем сильнее расхождение между реализованной нишей и потенциальной экологической нишей.
При жестких стрессовых воздействиях иногда возрастает значение коэффициентов вариации для многих морфоструктурных признаков. Но имеет место и противоположный процесс. Н.Г. Баш- товым (1991) сформулирована новая концепция о реализации популяциями и особями растений в условиях антропогенных нагрузок стратегии защиты, которая проявляется в повышении морфологической интегрированности особей, когда основные их признаки снижают уровень варьирования и получают большую скор- релированность. Для обеспечения защиты у растений осуществляется изменение аллокации веществ. Стратегия защиты реализуется на первых ступенях всех типов антропогенных нагрузок.
Экологический стресс в некоторых случаях существенно изменяет характер жизненной формы растения и их морфологическую структуру. Описано много таких случаев. У луговых дер- новинных злаков может полностью подавляться процесс кущения с образованием однопобеговых растений. Еще И.Г. Серебряков (1952) отмечал у клевера ползучего Trifolium repens превращение монокарпичных побегов в ноликарпичные. У рыхлокустового злака полевицы собачей Agrostis canina в неблагоприятных условиях при кущении побеги начинают формироваться ин- травагинально, то есть наблюдается переход к плотнокустовому типу кущения. В условиях низкой аэрации почвы у девясила британского Inula britannica перестает развиваться главный корень, и корневая система из стержневой превращается в мочковатую.
Среди изменений структуры популяций под действием стрессов внешне наиболее выражены, а часто и наиболее существенны, изменения численности особей в популяции (рис. 12.2). Такие снижения численности особей могут начинаться сразу с начала действия стрессора и быть плавными (кривая А), проявляться с задержкой (кривая Б) или после более продолжительной задержки в популяции могут сразу погибать практически все особи (кривая В).
Установлено, что под влиянием стрессовых факторов у большинства растений снижается продолжительность репродуктивного состояния, а иногда оно вообще выпадает, тогда как продолжительность пребывания растений в предгенеративном и по- стгенерэтивном состоянии увеличивается. В других случаях в зависимости от вида растения, жизненной формы и условий из популяций могут почти полностью выпадать либо предгенера- тивные, либо постгенеративные особи. Все это, естественно,
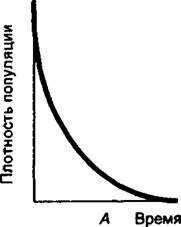
Рис. 12.2. Основные модели изменения плотности популяций растений при воздействии внутрипопуляционных факторов или внешних стрессоров (пояснения в тексте)

Рис. 12.2.Б
ведет к глубоким изменениям онтогенетических спектров популяций. Изменяются и виталитетные спектры популяций. Как правило, они приобретают депрессивный характер. Конечно, весь комплекс таких трансформаций специфичен и зависит от характера воздействующего фактора. И.М. Ермаковой и Н.С. Сугоркиной (1991) было установлено, что на лугах под влиянием стогования виды с разными стратегиями жизни изменяют структуру популяций по-разному.
200

Особый характер имеет такой все более распространяющийся стрессовый фактор, как рекреация - отдых населения в природных экосистемах, который сопровождается вытаптыванием, сбором красиво цветущих растений, организацией места отдыха

4 Scitla sibirica
-j Д Плотность S Фтомассз
100
120
4 Corydalis soiida
4 Anemone ranunculoides
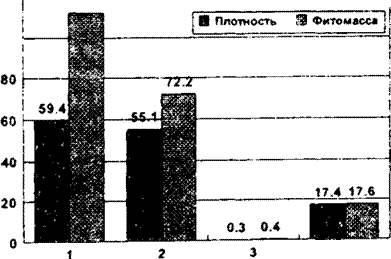
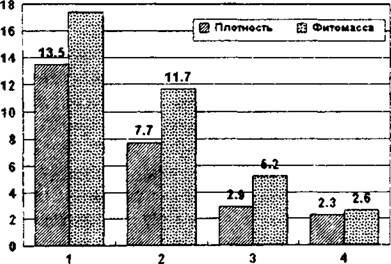
^uc. 12.3. Изменение плотности особей (шт./м2) и запаса фитомассы г/ м2) у четырех видов эфемероидов по нарастанию рекреационной
нагрузки (ступени 1-4)
и т.п. Рекреация ведет к измельчанию особей, в данном случае это универсальный эффект. Иногда рекреация оказывается для отдельных видов и положительным фактором: за счет снижения эдификаторного действия конкурирующих видов, менее устойчивых к рекреации, создаются более благоприятные условия для ассектаторов (Вахрамеева, 1991). Некоторые авторы считают рекреацию одним из наиболее мягких антропогенных факторов. Но это далеко не всегда так. Как видно из рис. 12.3, у четырех видов эфемероидов широколиственных лесов рекреация весьма существенно снижает плотность популяций и запас фитомассы растений.
Для устойчивого существования любой популяции в ней должно существовать определенное внутрипопуляционное разнообразие особей по полу, возрасту, онтогенетическому и вита- литетному состоянию. Компьютерное моделирование и обширный полевой материал, полученный разными исследователями, показывает, что разнообразие особей в популяции, находящейся в среде с разным уровнем стрессового воздействия, может быть сведено к пяти основным моделям (рис. 12.4). В местообитаниях, близких к оптимальным, с маловыраженной микромозаичностью распределение особей в репрезентативной выборке соответствует типу А. В этом случае в популяциях преобладают особи со средними значениями морфометрических показателей, а особей с отклонениями от этих показателей тем меньше, чем сильнее эти отклонения. Такой тип часто свойственен эдифика- торам. Тип А хорошо аппроксимируется нормальным статистическим распределением. Тип Б характерен для растений, произрастающих в местообитаниях с выраженной мозаичностью при примерно равной площади основных элементов мозаичности. Встречаемость особей разной размерности здесь приблизительно одинакова. Тип Б аппроксимируется либо нормальным распределением с низким значением эксцесса, либо равномерным статистическим распределением. Тип В характерен для растений контрастных местообитаний с наличием элементов мозаики, благоприятных для растений данного вида, и элементов, крайне неблагоприятных. На этой основе появляются двухвершинные и даже многовершинные типы распределения. Типы Г и Д свойственны популяциям растений, произрастающим в благоприятной и неблагоприятной среде соответственно. Распределение особей в статистических рядах в этих случаях асимметрично.

Рис. 12.4. Основные типы распределения особей растений по их размеру в популяциях растений по уровню их стрессового
Ингибирования (A)

(Б)

(Г)
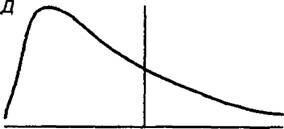
(Д)
При изменении условий среды популяция адаптируется к ним как целостная биосистема и постепенно стремится достичь состояния оптимального уровня внутрипопуляционного биоразнообразия. Схематически этот процесс представлен на рис. 12.5. Уровни разнообразия в популяциях ненарушенных природных систем наиболее близки к оптимальным значениям.
При постепенном снижении стабильности среды оптимальные значения внутрипопуляционного разнообразия могут увеличиваться, так что популяции, адаптированные к менее стабильным средам, приобретают более высокое внутрипопуляционное разнообразие. Увеличение внутрипопуляционного разнообразия с приближением его к оптимальному значению, расширение толерантности особей, рост рождаемости и снижение смертности повышают устойчивость таких популяций. Дальнейшее прогрессирующее уменьшение внутрипопуляционного разнообразия особей со все
Рис. 12.5. Соотношение между внутри популяционным разнообразием и жизнеспособностью популяции. Opt. - оптимальный уровень
разнообразия
большим отклонением условий произрастания от оптимума ведет к увеличению смертности, снижению рождаемости и обычно уменьшает общий запас фитомассы в популяции. Снижение, а также повышение внутреннего разнообразия биосистем, быстрые изменения среды ведут к снижению жизнеспособности популяций. Таким образом, внутрипопуляционное разнообразие во всех его формах нужно рассматривать как динамичный параметр, который лежит в основе стабильного существования популяций растений.
В.М. Остапко (2005) подразделял трансформации популяций под влиянием антропогенных факторов на три вида:
- прогрессивные, состоящие в усложнении структуры популяции;
- компенсационные, сохраняющие основные показатели структуры и функционирования за счет изменения второстепенных свойств;
- регрессивные, выражающиеся в упрощении, повышении однородности и унификации функций особей в популяции.
Особой формой антропогенного воздействия на популяции растений является радиоактивное загрязнение среды, которое происходит при использовании атомной энергии и связано с малыми или большими (типа Чернобыльской катастрофы) авариями на АЭС,
При поступлении и накоплении радионуклидов у относительно радиочувствительных видов растений в зависимости от физико-химических характеристик радиоактивных веществ, их количества и мест локализации могут наблюдаться те же радиобиологические эффекты, что и при эквивалентных поглощенных дозах внешнего облучения животными и человеком: радиационная стимуляция, морфологические изменения, ускорение старения и сокращение продолжительности жизни, деструкция ДНК, гибель.
Существование популяций в среде с повышенным радиационным фоном связано и с рядом дополнительных обстоятельств. Первое и основное из них - способность некоторых радиоактивных веществ избирательно накапливаться в отдельных тканях и органах растений, что приводит к относительно высоким локальным уровням облучения. Второе - увеличение опасности действия альфа- и бета-излучений, которые в малой степени влияют на растения в условиях внешнего облучения, но могут стать необычайно сильным источником ионизирующей радиации при поступлении внутрь клеток и клеточных структур. Третье обстоятельство вытекает из повышенной длительности сроков облучения. Ввиду специфики метаболизма растений по сравнению с животными радионуклиды, аккумулируясь в тканях, практически не выводятся из них. При огромных периодах полураспада ^Sr, 137Cs, 2зэри у многолетних растений эти радионуклиды будут создавать фон повышенного облучения в течение всего онтогенеза.
По мере увеличения продолжительности пребывания многолетних растений в зоне с радиационной аномалией следует ожидать селективного увеличения полиморфизма и физиологической нестабильности растений. В этом случае пролонгированное радиационное воздействие на меристемы конусов нарастания может способствовать глубокому преобразованию генетического и физиологического гомеостаза, что, по-видимому, способно привести к микроэволюционным сдвигам. Трансформация популяционной структуры растений с территорий радиоактивного заражения остается практически не изученной.
В целом, комплексный популяционный анализ с определением характера и степени стресс-трансформации отдельных особей и популяций дает богатый информационный материал и характеризует как состояние популяций, так и экосистем, в которых они находятся.
Еще по теме 12.3.2. Стрессоры и стресс:
- СТРЕСС. НЕВРОЗЫ. ЭПИЛЕПСИЯ. ЭКЛАМПСИЯ СТРЕСС - STRESS
- Биологические особенности кукурузы и ее устойчивость к абиотическим стрессорам
- 10.4. СТРЕССЫ В ЖИВОТНОВОДСТВЕ И МЕРЫ ИХ ПРОФИЛАКТИКИ
- Стресс и адаптация
- 5-11** Патологический стресс (дистресс)
- 5-11* Физиологический стресс (эвстресс)
- 5-11. Экспериментальная эволюция. Аршавский, доминанта и стресс
- 4* Вентребер, старец-пионер
- ЭКОЛОГИЯ И РАЗМНОЖЕНИЕ ЖИВОТНЫХ
- ЭКОЛОГИЯ И РАЗМНОЖЕНИЕ ЖИВОТНЫХ
- ВВЕДЕНИЕ
- ПРОФИЛАКТИКА АБОРТОВ
- ПРОФИЛАКТИКА АБОРТОВ
- А.А. Чиркин, И.М. Прищепа, А.Н. Дударев. основы ксенобиологии, 2003
- 11-3* Эволюция снизу
- АЛИМЕНТАРНОЕ БЕСПЛОДИЕ
- АЛИМЕНТАРНОЕ БЕСПЛОДИЕ
- НАРУШЕНИЯ ОБМЕНА ВЕЩЕСТВ Ожирение (Adipositas)
- 5-13* Поддержание нормы и ее смена