Распределение корма
У двух видов Bombus, запасающих пыльцу (В. terrestris и В. terricola), личинки находятся в общих ячейках (камерах) до последнего возраста. Рабочие особи проделывают в воске дырочку, чтобы проверить, как обстоят дела, а затем вновь ее заделывают; на последней личиночной стадии она остается открытой все время.
Промежутки между осмотрами (в среднем 11 мин) примерно вдвое короче, чем между кормежками (в среднем 20 мин). Кормление происходит нерегулярно, потому что рабочим особям часто приходится стоять в очереди за пыльцой у складов; это может занимать у них до 5 мин. Объясняется ли это недостатком складских помещений или самой пыльцы? Когда в одном эксперименте (Pendrel, Plowright, 1981) удалили часть рабочих особей, другие компенсировали это, повысив скорость кормления на 28%. Но если шмели способны на такие усилия, то можно полагать, что обычно они не слишком утруждают себя в гнезде.Подобное «обобществление» запасов пыльцы-явление необычное; даже пресоциальные пчелы распределяют свой расплод по нескольким ячейкам, снабженным комочками пыльцы. Эти комочки могут быть двух разных размеров; некоторые виды Evylaeus, как общественные, так и одиночные, откладывают неоплодотворенные яйца на маленькие комочки пыльцы, а оплодотворенные яйца-на большие, т. е. комочки, предназначенные самцам, меньше, чем комочки для самок (Knerer,
- . У принадлежащей к той же группе Lasioglossum zephyrum матка, как только она выделилась из группы равноправных самок, занимает в гнезде место, с которого она может оказывать влияние на деятельность рабочих особей, например в конце цепочки пчел у входа в гнездо или в месте разветвления тоннелей. Заняв такую позицию, она встречает и «подталкивает» рабочих. Она проводит также короткие осмотры ячеек, и Buckle (1982) высказал предположение, что она собирает информацию о ходе строительства ячеек, с тем чтобы направлять рабочих туда, где в них есть надобность. У Ceratinini, строящих камерные гнезда, пыльца не подвергается специальной обработке и не запасается, как у других пчел; смешав ее с жидкостью, отрыгиваемой из зоба (возможно, это нектар), рабочие помещают ее около личинок, которые кормятся все вместе. У других видов рабочие кладут кусочки корма прямо на тела личинок так, чтобы они могли дотянуться до него ротовыми частями.
У Ехопеига личинки, по-видимому, сигнализируют о том, что им нужен корм, движениями челюстей и головы; в ответ на это рабочие захватывают их своими челюстями и отрыгивают им в рот прозрачную жидкость (Sakagami, 1960). В линейных гнездах, устроенных в полых стеблях, личинки нередко бывают расположены очень упорядоченно: яйца откладываются в конце гнезда, противоположном входу, а личинки по мере развития продвигаются вперед и в конце концов окукливаются около входа (Michener, 1974). Такой порядок создается постепенно, он не устанавливается рабочими особями в результате прямой реакции на существование личинок разных классов.
У безжалой пчелы Melipona favosa строить ячейку начинают многочисленные пчелы-строительницы, но лишь несколько рабочих особей (от 5 до 8) заканчивают ее и снабжают кормом; в каждую ячейку кладется от 10 до 18 порций корма. Матка, находящаяся обычно на дне гнезда или в зоне запасов, время от времени инспектирует новые ячейки, и если обнаруживает ячейку с гладким воротничком, то начинает барабанить по ней. В ответ на это она редко получает корм, так что, возможно, это сигнал об откладке яиц. Непосредственно перед тем как отложить яйцо, матка поедает часть корма, предназначенного личинкам, а в дальнейшем съедает все кормовые яйца, если их откладывают рабочие особи, отделывающие ячейки и снабжающие их кормом (Sommeijer et al., 1982).
У медоносной пчелы весной не бывает недостатка в ячейках, поскольку зимой в улье нет расплода; однако матке приходится откладывать яйца только в вычищенные и выстланные внутри ячейки в той части улья, где рабочие особи способны поддерживать температуру на нужном уровне.
Эго, конечно, зависит от численности рабочих. Матка питается слюной, которую они вырабатывают из имеющихся в их организме резервов и из запасенной пыльцы; летом матку усиленно облизывают и скребут мандибулами. Пчела-кормилица, гипо(про)фарингеальные железы которой активно вырабатывают сйкрет, внимательно осматривает каждую ячейку, прежде чем поместить капельку жидкости на стенку ячейки или на личинку (по-видимому, она не удаляет из ячейки слюну или мочу). Если пища дается личинкам такими крошечными порциями, то неудивительно, что число «кормовых» посещений невероятно велико и одна пчела за всю свою жизнь может вырастить лишь двух или трех личинок. Не следует думать, что каждая кормилица занимается одной определенной группой ячеек,-она может носить корм многим сотням, если не тысячам личинок. Существует, однако, важная регуляторная особенность: когда корма мало, кормилицы снабжают им только обитателей немногих смежных ячеек (Gontarski, 1949), причем распределение корма целиком контролируется рабочими особями, так как личинки, отделенные от своих соседей стенкой ячейки, никак не могут конкурировать за корм.Хотя у пчел личинки не «кормят» рабочих особей, как это случается у ос, рабочие иногда обмениваются между собой пищей. Проситель вводит свой язычок в ротовую область потенциального донора, который выделяет капельку нектара, чего он не сделал бы без такого стимула (Free, 1959, р. 194). Без сомнения, донор, у которого зоб пуст, при первой возможности выходит из гнезда и отправляется фуражировать. Пыльцу, которую поедают пчелы-кормилицы, приносят сборщицы и кладут в подготовленные ячейки для временного хранения; сборщицы, которые не могут освободиться таким образом от груза, неохотно отправляются на дальнейшую фуражировку, так что по мере накопления пыльцы в гнезде интенсивность ее сбора снижается. Личинки стимулируют молодых пчел, чтобы те их кормили; при этом они используют главным образом тактильные стимулы, но частично также и обонятельные. Матки стимулируют сбор пыльцы даже в отсутствие расплода (Free, 1967; Free, Williams, 1971).
Семья муравья Amblyopone pallipes состоит всего лишь из одной самки и 9-16 рабочих особей. Откладка яиц происходит в августе, а выращиваемый из них расплод достигает зрелости в августе следующего года и дает как половых особей, так и рабочих. Таким образом, в каждый данный момент в семье преобладает расплод, находящийся на какой-то одной стадии развития; многолетний цикл развития у этого вида строго упорядочен и связан с временами года (Traniello, 1978). У Myrmicinae маленькие личинки получают отрыгиваемую рабочими жидкую пищу, а крупные - кусочки твердой пищи, либо животной, либо растительной (семена)
(Le Masne, 1963). Личинки первого возраста получают кормовые яйца, если они есть; личинкам второго возраста к этому добавляют отрыгиваемые соки, а личинкам третьего возраста-кусочки животной пищи. Третья личиночная стадия продолжается долго и включает два периода спячки, связанные с прекращением роста летом для подготовки к зимовке. Первый такой период блокирует рост личинок, развитие которых идет по «рабочему» пути, а второй-рост личинок, развивающихся в репродуктивных самок. Хотелось бы для простоты считать, что существуют личинки двух типов-дающие рабочих и дающие репродуктивных самок; однако последние на самом деле лабильны в отношении касты и могут переключаться обратно на путь развития рабочей особи. Зимой у Myrmica численное отношение рабочие / личинки варьирует около среднего 0,88, причем существует корреляция между числом личинок и числом рабочих и еще более тесная корреляция между их весами. Это обусловлено тем, что число рабочих особей определяет число яиц, откладываемых самкой, выживание этих яиц и вылупляющихся из них личинок, а также рост личинок; особенно это относится к личинкам третьего возраста, так как на этой стадии их вес увеличивается по меньшей мере втрое (Brian, 1965, 1979а). Все это так, когда корм имеется в достаточном количестве; если же количество корма ограничено, то его получает лишь небольшое число крупных личинок, а остальные личинки уничтожаются, обычно во время линьки.
Вероятно, наличие «голодных» рабочих особей с сильными хищническими наклонностями и слабых, не так легко распознаваемых рабочими личинок-достаточное условие для уничтожения последних.О I бор более крупных личинок и резервных куколок также целесообразен, поскольку они наиболее близки к зрелости и могут раньше начать помогать рабочим особям в гнезде, освобождая старших рабочих для охоты. Впавшие в спячку личинки третьего возраста могут продержаться без пищи довольно долгое время даже при температуре, подходящей для развития, но если они протянут слишком долго, то им придется вступить в конкуренцию с вновь вылупившимися весенними личинками, которым рабочие особи отдают предпочтение. В природных выводковых камерах личинки третьего возраста распределены достаточно свободно, каждая из них лежит на спинке и к каждой есть свободный доступ с потолка или со стен, кормят их довольно равномерно; в экспериментах, в которых личинок помещают на плоские пластинки, рабочие особи складывают их в кучки (вероятно, для того, чтобы их можно было окружить рабочими, обеспечивающими защиту), и хотя эту кучку постоянно обследуют и переворачивают, верхние личинки получают больше корма и растут быстрее. Таким образом, равномерность распределения корма, даже когда его вполне достаточно, зависит от устройства камеры, которая должна иметь скошенные стенки и скаты и обеспечивать физически прочную структуру, в которой рабочие особи могут расположить личинок, не опасаясь постороннего вторжения.
Иарядgt; с устройством гнезда присутствие самок тоже способствует равномерному распределению корма, возможно потому, что они вообще стимулируют его раздачу. Это не связано с тем, что они сами потребляют много пищи (тело мертвой самки оказывает почти такое же стимулирующее действие); это просто одно из проявлений катализирующего влияния самок на их семьи, повышающее эволюционную приспособленность семьи. У видов рода Myrmica личинки (рис. 10.1) ведут себя довольно сдержанно; обычно, если только они не слишком голодны, личинки ждут, пока им предложат корм, который они иногда принимают, а иногда отвергают; последнее, видимо, невыгодно для них, так как личинок, упорно не желающих брать корм, рабочие переводят в неактивную группу, т.
е. в группу, где находятся диапаузирующие личинки третьего возраста и личинки, переставшие принимать пищу незадолго до метаморфоза (предкуколкй); у первых «аппетит» может быть стимулирован вниманием со стороны рабочих особей. Хотя голова этих личинок привлекает внимание рабочих, а вытягивание головы и ее повороты из стороны в сторону служат сигналом, что личинка готова принять пищу, именно способность личинок брать пищу у кормилиц, съедать ее и затем снова брать заставляет кормилиц в свою очередь отправляться к фуражирам и просить у них корм (Brian, Abbott, 1977).Возможность побудить личинок к дальнейшему поглощению корма
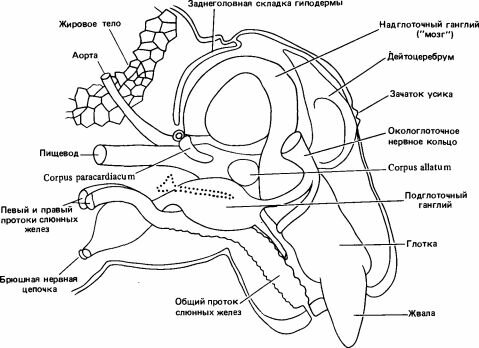
Рис. 10.1. Продольный разрез головы личинки Myrmica последнего возраста. (Weir, 1959.)
в сочетании со способностью самки стимулировать его распределение влияет на определяемое внутренними факторами «решение» о том, расти ли личинкам прямо до превращения в имаго или же диапаузировать и провести зиму на последней личиночной стадии. При температурах ниже 22°С рабочие отдают предпочтение личинкам, проявляющим явную тенденцию метаморфизировать; однако и многим менее склонным к метаморфозу личинкам удается получить достаточно корма, чтобы преодолеть первый физиологический барьер к росту-первичную диапаузу, и они завершают развитие, превращаясь в рабочих особей до начала зимы. Размеры остатка, как мы видели, зависят он численности и веса рабочих. Зимовка семьи, в которой имеются личинки, а не только взрослые особи,-это способ вместить в данный год еще один цикл развития потомства; в результате получаются два цикла-летний от яиц до взрослых особей и зимний, в котором личинки активны осенью и весной, а середину зимы проводят в спячке, находясь в третьем возрасте. Летний цикл дает рабочих особей, а зимний - репродуктивных особей и рабочих.
На следующий год новая партия яиц может начать развиваться раньше, чем личинки предыдущей партии достигнут зрелости; хотя при этом создается перекрывание и расплоды конкурируют за приносимые рабочими ресурсы, но зато рабочие особи получают возможность продлить период своей активности, захватив часть зимы,-до тех пор, пока личинки способны поглощать корм, а это они могут делать даже при понижении температуры примерно до 10°С (Brian, 1965).
При нос і роении модели сообщества Myrmica (Brian et al., 1981) было трудно включить в нее конкуренцию за корм между самками и личинками так, чтобы модель оставалась стабильной. С этой целью было введено предположение, что имеющийся корм распределяется между этими двумя категориями потребителей в соотношении, соответствующем весовому отношению между яйцом и куколкой, так что на каждую образовавшуюся куколку откладывается только по одному яйцу, которое должно ее заменить. Таким образом поддерживается непрерывный поток развивающихся единиц, проходящих через кормовую камеру, распределяется нагрузка на рабочих особей и обеспечивается постоянное наличие личинок, поскольку приток яиц и отток куколок происходят с одинаковой скоростью. Другой способ, позволяющий уменьшить колебания численности населения,-это запасание корма.
У Myrmica, после того как большинство личинок метаморфизировало, в пакетах яиц, отложенных самкой, оказывается много кормовых яиц. Поэтому в периоды, когда личинок мало, фуражиры могут продолжать сбор корма, а кормилицы перерабатывают его и запасают в виде желтка (Brian, 1977).
Еще по теме Распределение корма:
- Перевариваемость и усвоение корма консументами.
- КОРМА, ПЕРЕРАБОТКА КОРМОВ, ПИЩЕВАРЕНИЕ
- ОТЫСКИВАНИЕ ИСТОЧНИКОВ КОРМА. МОБИЛИЗАЦИЯ ПЧЕЛ НА ЕГО СБОР
- Свойства углеводного корма.
- Популяции насекомых при смене корма
- Значение отдельных видов корма
- Стадиальное распределение
- РАСПРЕДЕЛЕНИЕ МИКРООРГАНИЗМОВПО ПОЧВЕННОМУ ПРОФИЛЮИ ИХ ПЕРЕМЕЩЕНИЕ
- Горизонтальная структура - типы распределения и размещения особей
- Глава б. КОРМА, ТРОФИЧЕСКИЕ СВЯЗИ И ПИЩЕВОЕ ПОВЕДЕНИЕ
- Распределение и депонирование токсикантов в организме
- 5.5. КОРМА, ВЫЗЫВАЮЩИЕ ЗАБОЛЕВАНИЯ ВСЛЕДСТВИЕ НАЛИЧИЯ ТОКСИНОВ ЕСТЕСТВЕННОГО ПРОИСХОЖДЕНИЯ
- Распределение активности во времени суток
- Общие принципы распределения токсикантов в организме.
- Распределение жизни в биосфере.
- ПРОТОТИПИЧЕСКОЕ РАСПРЕДЕЛЕНИЕ АРХИТЕКТОНИЧЕСКИХ КОМПЛЕКСОВ В ОКЕАНСКОЙ ПЕЛАГИАЛИ
- Закономерности распределения гетеротрофных сообществ
- РАСПРЕДЕЛЕНИЕ ЭЛЕМЕНТОВ В РАЗЛИЧНЫХ КОМПОНЕНТАХ ЭКОСИСТЕМ СУШИ
- ЛАНДШАФТНО-БИОТОПИЧЕСКОЕ РАСПРЕДЕЛЕНИЕ КОМАРОВ
- Влияние морских течений на распределение температу р в водных массах.