Закономерности распределения гетеротрофных сообществ
В целом, доминирование разных трофических групп определяют концентрация взвеси и содержание органического углерода в верхнем слое осадка (рис. 3.18).
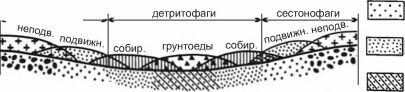
Во внутренних морях, лишённых глубоких котловин, возможно из-за большого потока детрита, достаточного для поддержания доминирования собирающих детритофагов, зона доминирования грунтоедов отсутствует.
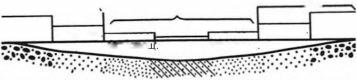
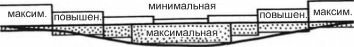
При чрезмерно большом осаждении детрита (например в зоне апвеллингов) может существенно (до нуля) падать концентрация кислорода в придонных слоях воды, макрозообентос сильно угнетён или вовсе отсутствует.При параллельных берегам изобатах и течениях трофические зоны располагаются, преимущественно, поясами в следующей последовательности: фильтраторы эпифауны - фильтраторы инфауны - собирающие - грунтоеды (рис. 3.19). Это чередование зон выражено чётко и обусловлено затуханием гидродинамики вод с увеличением глубины и уда-
гидродинамика
напряжённая
напряженная А_
ослабленная
максим
повышен
максим.
повышен
минимальная
затухаюі
затухающ
придонная взвесь
содержание Сорг в осадке и интенсивность осадконакопления
трофические зоны
сестонофаги
донные осадки
Рис. 3.19. Связь распределения трофических зон с рельефом и гидродинамикой. По
А.П. Кузнецову (1980).
гравиино-галечные и крупные пески
средние и мелкие пески
крупные и мелкие алевриты
лени ем от берега.
Если изобаты и течения проходят не строго параллельно берегам, поясное распределение зон может сменяться смешанным или даже пятнистым. Однако во всех случаях, даже когда общая картина распределения оказывается крайне сложной, как, например, в Баренцевом море (рис. 3.20 и 3.21), зоны фильтраторов располагаются в местах с повышенной гидродинамикой, более грубозернистыми осадками, повышенным содержанием взвеси в придонных слоях. Такие места расположены либо ближе к берегам, либо совпадают с выпуклостями рельефа. Зона потребителей детрита из грунта (собирающих и грунтоедов) приурочена к понижениям дна, более мелкозернистым осадками с повышенным содержанием органики, преобладанием процессов оседания взвеси над ее переносом.«В Баренцевом море районы активной гидродинамики располагаются в сферах главных потоков баренцевоморских течений (рис. 3.20). Это районы: Нордкап, Мурманское прибрежье, юго- восточное мелководье, Шпицбергенско-Медве- жинское мелководье и некоторые другие. Течения здесь имеют скорость до 25 см/сек (0,9 км/час) (Агеноров, 1946; Топорков, 1970). Районы эти выстланы песками и более грубозернистыми осадками и являются основными областями распространения неподвижных и подвижных сесто- нофагов (рис. 3.21). В то же время, районы максимально ослабленной гидродинамики совпадают с областями больших циклонических круговоротов, расположенных своими центрами над наибольшими понижениями дна (западный желоб, центральная и северо-восточная впадины и др.), выстланными мелкоалевритовыми илами и пелитами, обогащенными органикой и населенными фауной грунтоедов. Между первыми и вторыми располагаются области с затухающей гидродинамикой, где грунты — преимущественно крупные алевриты. В фауне преобладают собирающие детритофаги» (Кузнецов, 1980: 171).
Изучая распределение трофических зон на шельфах Мирового океана, А.А. Нейман (1988) показала, что на широких шельфах, где гидродинамические процессы и процессы осадконакопления протекают по замкнутому циклу, имеются зоны доминирования всех 4 типов.
Чем уже шельф, темittamp;
ЛҐ+Т+-
Рис. 3.20. Схема областей различной гидродинамической активности в Баренцевом море, выделенных по трофическому составу бентоса.
1 — активной гидродинамики; 2 — переходной; 3 — замедленной гидродинамики; 4 — поверхностные течения. По А.П. Кузнецову (1982).
Рис. 3.21. Трофические зоны бентоса Баренцева моря и образующие их сообщества.
1 — фильтраторов эпифауны; 2 — фильтраторов инфауны; 3 — собирающих детритофагов; 4 — грунтоедов. Разные сообщества показаны разными подстрочными символами. По А.П. Кузнецову (1982).
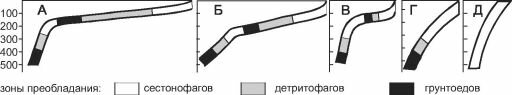
Рис. 3.22. Распределение трофических зон на шельфах разной крутизны и ширины.
А, Б — широкие шельфы с выраженной зоной осадконакопления в центральной части; В — узкий шельф со слабо выраженной зоной осадконакопления; Г, Д — увеличение крутизны шельфа приводит к сдвигу за пределы шельфа или к исчезновению зоны детритофагов. По А.А. Нейман (1988)
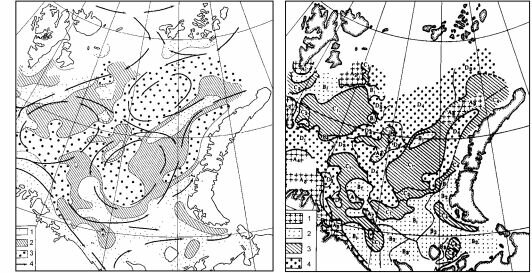
большая часть детрита выносится за его границы, поэтому на более узких шельфах развиваются не все зоны. Первой исчезает зона грунтоедов, затем детритофагов и даже фильтраторов инфауны (рис. 3.22). Ширина шельфа варьирует от 0 (некоторые места африканского побережья) до 1500 км (север Евразии и Северной Америки) (Баскаков, 1982; Баскаков, Шпайхер, 1982). Широкий шельф расположен преимущественно у берегов районов современного или плейстоценового оледенения: вдоль берегов Северного Ледовитого, севере и западе Тихого, севере и юго-западе Атлантического океанов, у Антарктиды, а также в Индо-Малайском архипелаге (рис. 3.23).
Существует еще одна, пока не получившая объяснения закономерность: на широких шельфах с хорошо развитой зоной седиментации, на самых тонких глинистых илах доминируют собирающие детритофаги, глотающие же преобладают на несколько более грубых илах.
На более узких шельфах, где зона седиментации не столь обширна, наоборот, на самых тонких илах доминируют не собирающие, а глотающие детритофаги.Описанная картина распределения трофических зон наблюдается в открытых частях шельфов. Выше, в самых прибрежных районах наблюдается отдельный набор трофических зон, к которому добавляется зона первичных продуцентов: макрофитов, мангров, гер- матипных кораллов.
Ниже шельфа располагается ещё несколько наборов трофических зон. Один набор начинается на кромке шельфа, где развивается зона фильтраторов эпифауны. Для неё очень характерны рифы агерматипных кораллов, заросли которых достигают в высоту (от уровня дна) нескольких десятков метров (до 35 м и даже более). Также для неё характерны зоны развития Porifera и Brachiopoda. Баренцевоморские сообщества губок и брахио-
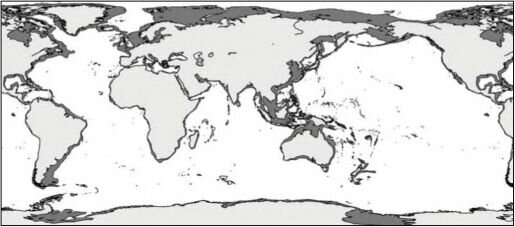
Рис. 3.23. Ширина шельфа в разных районах Мирового океана.
I
¦ ' —2 ИЗ [П]4 Ш5 Вб
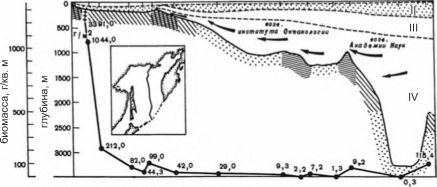
Рис. 3.24. Чередование трофических зон в Охотском море.
1,2 — фильтраторы эпифауны (1 — мелководные, 2 — глубоководные); 3 — фильтраторы инфауны; 4 — собирающие детритофаги; 5 — грунтоеды; 6 — потоки глубинных океанских вод. Римские цифры — разные водные массы: I — поверхностная, II — подповерхностная, III — главный термоклин, IV — глубинная. Кривая — средняя биомасса. По А.П. Кузнецову (1982).
под принадлежат к этой зоне. Другой набор начинается на переходе океанического ложа в ультраабиссальные желоба (рис. 3.24).
Характерная особенность океанического ложа — наличие больших районов с очень низким потоком детрита. Исследуя это явление, М.Н. Соколова (1977 и более ранние работы) разделила океаническое ложе на эвтрофные и олиготрофные районы. Олиготрофные районы расположены в центрах океанических круговоротов (рис.
3.25). Различие эв- трофных и олиготрофных районов обусловлено различием потоков детрита: в централь-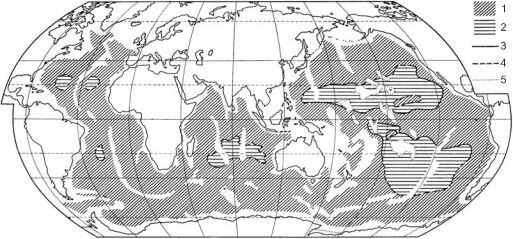
Рис. 3.25. Глубоководные донные трофические области (в пределах глубин более 3000 м).
1 — эвтрофные; 2 — олиготрофные; 3-4 — границы между ними: проведенные по весовому преобладанию де- тритоедов в траловых уловах с учетом распространения основных систематических групп детритоедов в макробентосе, весовой характеристики макро- и мейобентоса, а также условий накопления и преобразования органического вещества в поверхностном слое осадков (3); то же, но обеспеченные малым материалом (4); 5 — внешняя граница аккумулятивных шлейфов у островов и подводных горных сооружений. По Н.Г. Соколовой (1977).
ных частях океанов очень низкая первичная продукция на поверхности; в периферических она выше и к тому же детрит поступает и из неритических районов. Это различие приводит не только к падению биомассы в центральных районах океанов, но и к перестройке трофической структуры сообществ. В олиготрофных районах очень мало грунтое- дов и собирающих, и ведущая роль опять переходит к фильтраторам, но уже фильтрато- рам эпифауны рыхлых грунтов (рис. 3.26). Перестройка трофической структуры приводит к фаунистическим различиям: для олиготрофных районов не характерны (обычно вовсе в них отсутствуют) такие группы, как неправильные морские ежи, Isopoda, Scaphopoda, ас- цидии, многие семейства полихет (например, Ampharetidae, Terebellidae), морских звёзд, голотурий. Вместе с тем, таксонов, специфичных для олиготрофных районов, нет: все виды, встречающиеся в них, обитают и в эвтрофных районах.
Существенно, что различия трофической структуры характерны только для макро- бентоса, тогда как мейобентос эвтрофных и олиготрофных районов различается гораздо меньше.
В некоторых районах, например на вершинах подводных гор (Парии и др., 1993), были обнаружены бентосные сообщества, до 99,9% биомассы которых образовывали хищные животные: омары и другие крупные десятиногие ракообразные, хищные морские звёзды и ежи.
Изучение их рациона по содержимому желудка показало, что стандартные орудия сбора бентосных проб не учитывали их жертв: макропланктон и нектобентос. Наличие сообществ подобного типа дополнительно свидетельствует о том, что в состав бентосных сообществ следует включать и организмы, обитающие в придонных слоях воды (бентопелагиали), биология которых тесно связана с дном.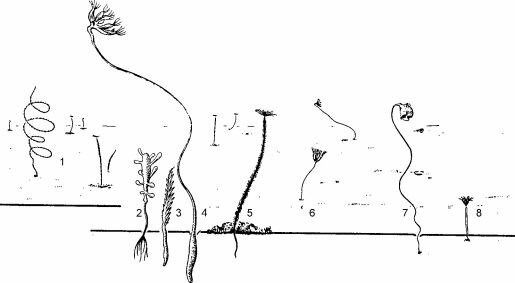
Рис. 3.26. Сообщество фильтраторов эпифауны рыхлых грунтов.
1 — Radicipes verrillii; 2 — Chondrocladia gigantea; 3 — Povanaria; 4 — Umbelula; 5 — полихета Potarnilla syrn- biotica; 6 — Cryptospongia enigrnatica; 1 — асцидия Culeolus; 8 — Kinetoskias. По А.И. Савилову (1961).
Автотрофные сообщества
В море существует три основных типа автотрофных сообществ макробентоса: макрофитов, мелководных рифообразующих кораллов с симбиотическими водорослями[†††††] и глубоководных гидротерм.
Еще по теме Закономерности распределения гетеротрофных сообществ:
- Гетеротрофные сообщества
- Общие закономерности вертикальной стратификации микробных сообществ
- Биогеоценозы гетеротрофного типа
- Продуктивность сообществ и жизненные стратегии водорослей. Морфофункциональный подход к изучению сообществ макрофитов
- СОСТАВ СООБЩЕСТВ И КОМПЛЕКСЫ СООБЩЕСТВ
- ПРОТОТИПИЧЕСКОЕ РАСПРЕДЕЛЕНИЕ АРХИТЕКТОНИЧЕСКИХ КОМПЛЕКСОВ В ОКЕАНСКОЙ ПЕЛАГИАЛИ
- Распределение активности во времени суток
- РАСПРЕДЕЛЕНИЕ МИКРООРГАНИЗМОВПО ПОЧВЕННОМУ ПРОФИЛЮИ ИХ ПЕРЕМЕЩЕНИЕ
- Горизонтальная структура - типы распределения и размещения особей
- РАСПРЕДЕЛЕНИЕ ЭЛЕМЕНТОВ В РАЗЛИЧНЫХ КОМПОНЕНТАХ ЭКОСИСТЕМ СУШИ
- Рифовые сообщества Особенности рифовых сообществ
- Распределение и депонирование токсикантов в организме
- Общие принципы распределения токсикантов в организме.
- Распределение жизни в биосфере.
- Стадиальное распределение