Сообщества морских макрофитов[‡‡‡‡‡]
Морские макрофиты населяют диапазон глубин от супралиторали (зона заплеска) до 20-30 м, хотя известны и гораздо более глубоководные нахождения живых макрофитов в местах с очень высокой прозрачностью воды (более 100 и даже 260 м) (Возжинская и др., 1992; Littler et al., 1989).
Макроводоросли занимают в основном биотопы с твёрдым субстратом (скалы, валунные россыпи, ракушечники), а цветковые растения — с рыхлыми грунтами (пески, заиленные пески), хотя известны и исключения из этого правила : например, некоторые зелёные водоросли (Caulerpa, Chara, Udotea, Penicdlus) вегетируют на песках, а цветковое растение Phyllospadix — на скалах и валунных россыпях. Макроводоросли образуют также неприкреплённые скопления, например, саргассы Саргассового моря или филлофора Филлофорного поля Зёрнова. Положение нижней границы распространения макрофитов обусловлено минимально необходимым уровнем освещённости и наличием подходящего субстрата.При всём разнообразии, для зарослей макрофитов характерны:
- высокая продуктивность, сравнимая или даже превосходящая продуктивность сельскохозяйственных культур и тропических лесов (рис. 3.48);
- заросли макрофитов служат убежищем для постоянных и временных обитателей, многие из которых имеют коммерческое значение;
- талломы (листья) макрофитов замедляют движения воды, в результате их заросли становятся ловушкой для детрита (особенно это характерно для морских трав), что приводит к существенному увеличению роли детритофагов и даже грунтоедов.
Как субстрат, макрофиты гораздо сложнее субстрата-грунта, поскольку это не инертный субстрат, а компонент сообщества, меняющийся во времени. Он включён в трофические цепи этого сообщества. До половины органического вещества, синтезированного макрофитами, тут же выделяется в виде растворённой органики.
Основные блоки сообществ макрофитов это: (1) зарослеобразующие макрофиты; (2) их эпифитон и (3) эпи- и инфауна субстрата, к которому прикреплены макрофиты.
По трофической структуре зооэпифитон можно разделить на 2 группы: фитофаги и сестонофаги. Фитофаги питаются растениями, но, в отличие от наземных сообществ, в основном, не зарослеобразующим макрофитом, а его эпифитами, как многоклеточными, так и одноклеточными. Сестонофаги просто используют макрофиты в качестве субстрата, поднимающего их над дном, хотя и среди них есть виды, предпочитающие те или иные макрофиты. Способы прикрепления эпифитона весьма разнообразны (рис. 3.27).
Виды беспозвоночных, характерные для одного вида макрофита, исключительно редки, хотя некоторое предпочтение и может оказываться. Так, в Чёрном море на цисто- зире преобладают ракообразные и черви, на филлофоре — моллюски и мшанки, на руп- пии — полихеты и гастроподы, на зостере — бокоплавы (Макавеева, 1979). Одни и те же виды обитают на разных макрофитах, и на их распределение в большей степени влияют абиотические факторы (глубина, прибойность), нежели субстрат-макрофит. Этим сообщества морских макрофитов отличаются от сообществ наземных. Различие животного насе-
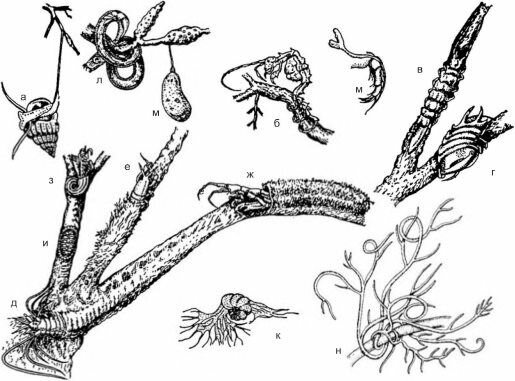
Рис. 3.27. Способы прикрепления эпифитона к макрофитам, а — нитью из слизи (Rissoa splendida), б-г — конечностями (б — Caprella acanthi/era, в — Synisorna capito, г — Naesa bidentata), д-ж — трубками: из слизи (д —Platynereis dumerillii), обрывков макрофитов (е —Leptochelia sa- vignyi), детрита и кусочков макрофитов (ж — Erichthonius difformis), з — известковыми домиками (Spirorbis pu- silla). и — раковиной (Lepidochitona cinerea), к — псевдоподиями (Cribroelphidium rnartcobi), л — секретом хвостовых желез (Cyatholaimus demani), м — секретом паутинных желез (Sclerochilus gewemulleri), н — ризоидами (Chondria califronica).Uo Е.Б. Макавеевой (1989) и Abbott, Hollenberg (1976).
ления разных видов макрофитов в первую очередь обусловлено геометрией таллома. При равных условиях, на более разветвлённых макрофитах выше видовое разнообразие, биомасса и численность.
Так, в Красном море численность эпифитона на сильно разветвлённой бурой водоросле цистозире, менее разветвлённой халимеде и лентовидной морской траве посидонии соотносится как 96:4:1, а биомасса — 13:2,6:1 (Макавеева, 1979).Что касается беспозвоночных, обитающих в грунте, под зарослями макрофитов, то специфических видов тут также нет, всё это — обычные верхнесублиторальные виды.
Классификация жизненных форм морских водорослей
Кроме общеботанической системы К. Раункиера, существует несколько классификаций жизеннных форм водных растений. Den Hartog и Segal (1964), модифицировав систему жизненных форм F. Clements (1920) (обе ссылки — цит. по: Raven, Richardson, 1986), предложили разделить их на две группы: прикреплённые к субстрату (attached plants) и неприкреплённые, или планофиты (planophytes). Прикреплённые растения подразделяют на гаптофиты (haptophytes), которые крепятся к твёрдому субстрату (валуны, скалы), размеры частиц которого превосходят само растение, и на ризофиты (rhizophytes), укоренённые или погружённые в рыхлый субстрат (песок, ил), частицы которого существенно меньше растения. К планофитам относят планктофиты (planktophytes, мелкие планктонные водоросли) и плейстофиты (pleustophytes, крупные плавающие или свободно лежащие на дне растения). Плейстофиты подразделяют на бенто-, мезо- и акроплейстофиты. Первые лежат на дне (пластообразующая Phyllophora, ряд фукоидов в солёных маршах и эстуариях), вторые плавают в толще воды, третьи — на её поверхности. Саргассы в Сар- гассовом море объединяют черты второй и третьей групп, поскольку встречаются и на поверхности, и в погружённом состоянии. Пример «чистых» акроплейстофитов — пресноводные ряска (Lemna) и хорошо известная аквариумистам пистия (Pistia).
Гаптофиты делят на эпилитные (растущие на камнях и скалах макроводоросли и морская трава Phyllospadix) и эпифитные (растущие на других растениях). В отдельную группу эпипсаммона объединяют микроскопические водоросли (диатомеи, цианобактерии и др.), которые прикрепляются к песчинкам, как к твёрдому субстрату, превосходящему их по размерам.
Жизненная форма ризофитов представлена морскими травами, рядом зелёных водорослей (Chara, Lamprotamnium, Caulerpa), а также карликовыми неприкреплёнными фукоидами.Den Hartog (1959) дал классификацию, построенную на морфологическом подходе. Исследуя морскую растительность Нидерландов, он выделил 14 типов жизненных форм: тип Laminaria, тип Fucus, тип Cystoseira, тип Desmarestia, корковые водоросли (тип Fdil- denbradtia), простые нитчатые (тип Ulothrix), разветвлённые (древовидные) нитчатые (тип Cladophora), трубчатые (тип Enteromorpha), пластинчатые (тип Ulva), кустистые (тип Chondrus), студенистые (тип Nemalion), известковые (тип Corallina), подушковидные (тип Vaucheria) и, наконец, группа микроскопических водорослей, таллом которых меньше 2 мм, в которую входят многие эпифиты, эндофиты и паразиты.
К.М. Петров (2004) предложил классификацию жизненных форм, основанную на внешнем облике и размерах донных растений: 1) Gramminida — морские травы; 2) Mag- noalgosa — водоросли с длиной таллома 1-4 м (крупные бурые водоросли); 3) Mesoalgo- sa — водоросли со слоевищами различной морфологии длиной 10-30 см; 4) Parvoalgo- sa — водоросли длиной 1-3 см (дерновино-образующие); 5) Corallida — красные известковые кустистые водоросли, длина талломов 2-4 см; 6) Crustida — корковые водоросли; 7) Epiphyton — эпифитные водоросли. Эти названия он предложил использовать при описании структуры донных фитоценозов.
Морфология многих водорослей-макрофитов весьма изменчива, многие способны формировать экологические формы — экады.
Еще по теме Сообщества морских макрофитов[‡‡‡‡‡]:
- Продуктивность сообществ и жизненные стратегии водорослей. Морфофункциональный подход к изучению сообществ макрофитов
- Классификация морских донных растительных сообществ
- Макрофиты
- Сукцессии и взаимоотношения макрофитов
- СОСТАВ СООБЩЕСТВ И КОМПЛЕКСЫ СООБЩЕСТВ
- Макрофиты[††††]
- Макрофиты — вредные вселенцы
- Рифовые сообщества Особенности рифовых сообществ
- Морские перья
- Морские звезды
- РОД КАЛИФОРНИЙСКИЕ МОРСКИЕ ЛЬВЫ GENUS ZALOPHUS
- Растительность морских побережий
- МОРСКИЕ ОХОТНИКИ