Стратегии жизненных циклов
Стремление обобщить накопленную информацию по аутоэкологии видов привело к возникновению концепций стратегий жизненных циклов (или просто жизненных стратегий), называемых также эколого-ценотическими стратегиями.
В конечном итоге, во всех концепциях рассматривают распределение энергии между различными популяционными процессами (миграции, особенности размножения) и особенностями биологии отдельных особей (рост, размеры, продолжительность жизни и т. п.).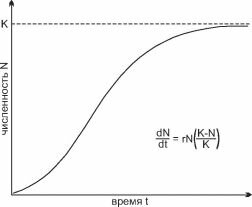
Рис. 2.22. Логистическая кривая роста численности популяции.
К — предельная численность вида, г — константа, отражающая максимально возможную скорость роста популяции.
Различные концепции обращают внимание на распределение аутэкологических характеристик по различным осям, которые авторы концепций считают наиболее значимыми.
Классификация, рассматривающая две жизненные стратегии — г-К
Самая простая классификация жизненных стратегий выделяет два основных типа. Названия этих типов различны. В первом варианте (McLeod, 1884, цит. по: Миркин и др., 2002) их назвали «капиталистами» и «пролетариями», сейчас их называют К- и r-стратегами (MacArthur, Wilson, 1967; Пи- анка, 1981; Ильяш и др., 2003) по коэффициэнтам логистического уравнения роста численности популяции г и К (рис. 2.22). Согласно одной из теорий (весьма популярной в современной экологии) и отдельным примерам, в сообществах с большим числом видов конкуренцию считают основным фактором отбора в условиях дефицита ресурсов, дефицит ресурсов же считают непременным свойством природных сообществ. Иными словами, конкуренцию за ресурс, согласно этим представлениям, считают основным фактором взаимодействия видов. Для многих экосистем или гильдий (например, сосудистых растений в наземных экосистемах или фитопланктона) это вполне допустимое упрощение.
Анализируя логистическую модель роста численности популяции, MacArthur и Wilson (1967) предположили, что в стабильных условиях конкуренция за ресурс приводит к отбору видов, у которых основной поток энергии через популяцию направлен на продуцирование более конкурентоспособных потомков (К-стратегия). Но большие затраты на производство отдельного потомка приводят к ограничению числа потомков, продуцируемых половозрелой особью. Это приводит к снижению максимальной скорости роста и снижению расселительных способностей вида. В экосистемах, где численность популяции ограничивают не лимитирующие факторы, за которые ведётся конкурентная борьба, а внешние воздействия (пресс хищников, постоянно действующие нарушения и т. п.), преимущество получают виды с более высокой скоростью размножения (r-стратеги). Эти виды производят большое количество потомков, на производство каждого из которых затрачена гораздо меньшая энергия. Напряжённость межвидовой конкуренции и связанная с ней степень доступности лимитирующего ресурса определяют положение видов на оси г- и К- стратегий. При К-стратегии виды-капиталисты направляют основной поток энергии на поддержание жизнедеятельности взрослых особей, при r-стратегии виды-пролетарии направляют основной поток энергии на размножение.Э. Пианка (1970, 1981) предположил, что в слабо нарушаемых сообществах преобладают К-виды, а в сильно нарушаемых — r-виды. С биологической точки зрения очень важна причина нарушений: являются ли они естественными, свойственными самим экосистемам, или их биотопам или же это антропогенные воздействия. К естественным нарушениям, в отличие от антропогенных, виды эволюционно адаптированы. Однако этот вопрос в данной теории не затрагивается.
R- и К-стратегии — крайние варианты, в реальных же условиях наблюдается континуум переходных форм.
На практике для определения положения видов на оси r-К используют различные показатели. Часто указывают, что для r-стратегов (их называют также оппортунистами) характерен короткий жизненный цикл с единственным периодом размножения в раннем возрасте и большой расход энергии на репродуктивные усилия.
Их считают характерными для эфемерных биотопов. Наоборот, для К-стратегов считают характерной долгую жизнь, многократное размножение, начинающееся сравнительно поздно. Но при изучении характеристик реальных видов эти показатели нередко дают противоречивую картину. Так, для подавляющего большинства видов морского макробентоса, а также многих рыб и головоногих, на суше — для деревьев, характерны высокая плодовитость, большой расход энергии на репродуктивные усилия, нередко сочетающиеся с большой продолжительностью жизни, причём плодовитость растёт с возрастом и размером. Более того, известны случаи, когда разные полы одного вида придерживаются разных стратегий или даже в пределах одного пола разные особи придерживаются разных стратегий размножения, это показано, например, для головоногих моллюсков (Лаптиховский, 2006). Поэтому при попытках отнесения к г-К-стратегам не отдельных специально подобранных видов, а достаточно полного набора видов из одной экосистемы обычно приходится говорить о свойствах г-К-статегов, при этом большую часть видов, часто, не удаётся отнести ни к тем, ни к другим. Тем не менее, г-К-классификация пользуется большой популярностью, в первую очередь из-за кажущейся её простоты и логичности. Трудности в практическом применении этой классификации возникают только при углублённом изучении биологии достаточно полного состава видов из одной экосистемы.Еще по теме Стратегии жизненных циклов:
- Классификации, рассматривающие три жизненные стратегии
- Продуктивность сообществ и жизненные стратегии водорослей. Морфофункциональный подход к изучению сообществ макрофитов
- R-K-стратеги и ценофобные-ценофильные виды
- 3. 2. Типы экологических стратегий
- Эколого-ценотические стратегии вилов и популяций растений
- Стратегия фуражировки
- ТЕХНОЛОГИЧЕСКАЯ СТРАТЕГИЯ
- Стратегии инвестиций азота
- Моя стратегия защиты
- 1.2. СТРАТЕГИЯ ЖИЗНИ. ПРИСПОСОБЛЕНИЕ, ПРОГРЕСС, ЭНЕРГЕТИЧЕСКОЕ И ИНФОРМАЦИОННОЕ ОБЕСПЕЧЕНИЕ
- ГЛАВА 8 Стратегии для городов и деревень
- Жизненные формы
- ЭВОЛЮЦИЯ ЖИЗНЕННЫХ ФОРМ
- 2.3. Жизненные формы организмов
- ЖИЗНЕННЫЕ ФОРМЫ У СПОРОВЫХ РАСТЕНИЙ
- Жизненные формы растений.
- Пятая динамика. Жизненные формы