Стратегии инвестиций азота
Экономика инвестиций азота — одно из краеугольных положений функциональной экологии растений. Успех вида в изменяющихся внешних условиях определяется тем, сколько азота, где и на какое время было введено в действие.
Ткани с высоким содержанием азота (протеина) отличаются высокой метаболической активностью (фотосинтез, дыхание, образование новых тканей; рис. 13.19), но одновременно по тем же самым причинам они очень привлекательны для фитофагов. Преждевременные потери таких растительных тканей из- за нарушений довольно тяжелы. Связи между максимальной степенью фотосинтеза Апих и содержанием азота на единицу листовой поверхности в пределах определенных мор- фотипов листа настолько тесные (и линейные), что по данным содержания азота с относительно малой вероятностью ошибки можно предсказать Длах (рис. 13.20). Благодаря тесному и также линейному соотношению между Атах и gmax (максимальная устьичная диффузионная проводимость для водяного пара; см. рис 13.14) выявляется двойная зависимость.Значения обеспечения азотом на единицу листовой поверхности также тесно коррелируют с SLA или образной величиной LIVLA (англ. leaf mass per area — масса листьев на единицу площади, г-м 2) (рис. 13.21). Листья, 1 г сухого вещества которых приходится на небольшую поверхность, содержат меньше N и больше С, поэтому для травоядных они менее привлекательны, но они также меньше ассимилируют и относительно инвестиций углерода на единицу поверхности довольно «дорогие».
Рис. 13.19. Распределение азота в листьях кроны Eucalyptus grandis (по R. Leuning et at). Зонирование имеет следующие основания: вверху (соответственно, снаружи) доступно больше света, «солнечные» листья имеют большую емкость фотосинтеза и поэтому больше протеина.
Листья внутренней части кроны получают меньше света, они старше, поэтому часто и более склероморфные (азот «разбавляется» за счет большего количества углерода), у них меньше площадь (SLA) и меньше потребность в RubisCo (ключевой фермент фотосинтеза)Очевидно, что такие различия в инвестициях могут быть компенсированы только длительностью функционирования. Листья с малым процентом азота и большим углерода должны долго оставаться актив-
Содержание азота в листьях, %
Между содержанием азота на единицу листовой поверхности и наиболее высокой относительно листовой поверхности степенью фотосинтеза при нормальном содержании С02 в воздухе существует тесная линейная связь. Угол наклона прямой, выражающей зависимость, снижается с уменьшением листовой поверхности на грамм сухого веса (SLA, м2- кг~’)- Листья с маленьким SLA, как правило, толстые и (или) более жесткие и долголетние. В них связано относительно большее количество углерода в нефотосинтетических структурах
30
\
о; 4

100 200 Масса листа на единицу поверхности, г -
Рис. 13.21. Зависимости между инвестициями азота и углерода в листья (по Ch.Korner).
ными, чтобы при низкой степени фотосинтеза возместить собственные затраты углерода и иметь возможность доставлять ассимиляты по всему растению. Такие листья обычно существуют долго (склерофиль- ные листья, вечнозеленые иглы хвойных).
В пределах определенных морфотипов листа существует линейная связь между количеством азота на единицу листовой поверхности и сухим веществом на единицу листовой поверхности (leaf mass per area, LMA= 1/SLA), независимо от вида растения. Такое группирование вдоль дискретных регрессий соответствует примерно одинаковой длительности жизни этих типов листьев (сходные концентрации азота в % от сухого веса и длительность амортизации, т.
е. времени, за которое лист оправдывает «издержки своей конструкции»). Снижение процентного значения азота с увеличением длительности жизни листа может иметь две причины: редукция содержания N в протопласте или увеличение массы клеточных стенок при таком же снабжении протопласта азотом. Как правило, обе эти причины имеют место300
Напротив, листья с большим SLA и высокой концентрацией N в % (см. табл. 13.1 в разделе 13.6.2) живут недолго, уже за несколько дней «амортизируются» (травянистые растения). Итак, по-разному снабжаемые листья после естественного старения разлагаются как подстилка с очень различной скоростью. Тип листа определяет таким образом и скорость рециклизации азота в системе. Отсюда вытекает многомерная взаимозависимость фотосинтеза, водного режима, длительности функционирования, риска поедания животными, разложения подстилки и круговорота азота в экосистеме (рис. 13.22).
Некоторые из этих зависимостей настолько прочны, что они значимы во всех биомах и для всех жизненных форм, как показал П.Райх на многих примерах (рис. 13.23). Как гласит основное правило, с возрастанием зрелости сообщества в сук- цессионном ряду чаще попадаются многолетние, богатые углеродом и более слабые в отношении ассимиляции типы листьев, если не существует сезонных огра-
ничений (листопадные виды в области с холодной зимой). Чем дольше питательные вещества связываются листьями, тем меньше риск потери их системой. Каждый листопад означает риск выноса из системы этих питательных веществ. Ограничения также устанавливаются и самозатенением растений. Длительное функционирование листьев часто в целом связано с медленным ростом или низким LAT.
Длительность

Разложение
подстилки
Круговорот азота -в экосистеме
Рис.
13.22. Центральная роль азота в особенностях листьевКонцентрация азота в листьях сама по себе препятствует разнообразным изменениям листьев или обусловливает их как в самом листе и отдельном растении, так и в экосистеме. Здесь даны некоторые особенно важные связи. Стрелки показывают направление воздействия, знаки «+» или «-» обозначают направление реакции в случае, если концентрация азота повышается (!) и интенсивность роста растения возрастает. Знаки «+» или «-» меняются на обратные, если концентрация N и связанная с ним интенсивность роста падают, причем отношение к последней проявляется только в пределах определенных морфотипов, что подтверждается на примере одинаково быстрорастущих летне-зеленых и вечнозеленых древесных растений. Часто принимаемое влияние степени транспирации на питание листьев не доказано и невозможно, поэтому в данном случае стрелка отсутствует. Высокая влажность почвы может способствовать как доступности азота, так и транспирации, но это — совпадение (т. е. первое никак не причина второго). Растения при постоянно насыщенном водяными парами воздухе прекрасно процветают без транспирации (движение воды и транспорт питательных веществ в ксилеме, тесно связанные с флоэмными функциями)
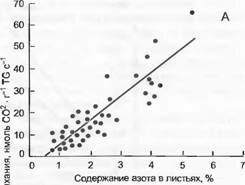
Рис. 13.23. Митохондриальное дыхание и концентрация азота в листьях (по Р. Reich et al.):
А — с увеличением концентрации азота в листьях увеличивается их темновое дыхание (ср. рис. 13.20); В— с увеличением темнового дыхания падает продолжительность жизни листьев. Эти сравнения охватывают листья растений всех климатических зон. Каждая точка обозначает отдельный вид
Молодые (рудеральные) сообщества растений состоят из быстрорастущих малолетних растений с богатыми азотом, тонкими, быстро амортизирующимися листьями, которые после отмирания очень быстро разлагаются. Некоторые растения с долго живущими листьями (например, хвойные деревья) сохраняют свои старые листья еще какое-то время в кроне, когда они из-за затенения более молодыми по сути дела больше не поставляют сколько-нибудь значимой нетто-продукции углерода.
На этой стадии такие листья представляют собой живые консервы питательных веществ, которые при необходимости могут быть извлечены (например, при весеннем распускании, потерях листьев из-за фитофагов, дефиците питательных веществ из-за высыхания верхнего слоя почвы). У ели (Picea abies) хвоинки старше 4 — 5 лет относятся к этой категории.До некоторой степени описанные явления дополняются другими защитными механизмами. Растения несомненно могут сдерживать определенных фитофагов путем синтеза алкалоидов, гликозидов, фенолов и терпенов, образования масел, млечного сока или смолы. Правда, эти защитные механизмы требуют затрат, и фитофаги часто к ним устойчивы. Смертельно ядовитый для человека и крупного рогатого скота тисс (Taxus baccata) для некоторых диких животных — излюбленное лакомство. Эвкалипты, несмотря на очень высокое содержание эфирных масел, ежегодно теряют до половины своих молодых побегов из-за фитофагов. Насколько известно, ни одна трава не является слишком ядовитой для улиток. Бедность протеинами и жестколистность представляются наиболее действенным средством для увеличения длительности жизни листьев. Чистая продуктивность недолговечных листьев с высокой активностью и долгоживущих с низкой может быть почти одинаковой, что объясняет, почему растения с комплементарными стратегиями листьев могут весьма успешно сосуществовать без принципиальных различий в степени роста и процветания. Примеры — кедровая сосна и лиственница (Pinus cembra и Larix decidua) в Альпах, черника и брусника (Vaccinium myrtillus и Vaccinium vitis-idaea) в березовой лесотундре Северной Европы или сезонно-зеленые и вечнозеленые виды в средиземноморском мелколесье.
Близко к понятию эффективности использования воды (water use efficiency, см. 13.5.4) широко употребляемое и столь же двусмысленные понятие эффективности использования азота (nitrogen use efficiency — NUE). NUE трактуется очень неоднозначно, часто для представления о том, насколько растение «эффективно», в качестве синонима NUE используется отношение N/С или просто % N, если растение обходится малым количеством азота.
При этом NUE может приводиться только для листьев или для содержания азота во всем растении. NUE определяется фотосинтезом относительно Атах, чаще всего без учета длительности жизни листьев и тем самым без учета «фотосинтетического урожая» за весь период их функционирования. Так как остается неясным, что же подразумевается в действительности под эффективностью (эффективны для чего?), этого понятия следует избегать и в крайнем случае заменять его «коэффициентом использования азота». Как правило, достаточно содержательно просто понятие концентрации азота (без какой-либо надуманной целесообразности).Как было объяснено в разделе 12.1, в естественных растительных сообществах, приспособленных к определенным местообитаниям и развивавшихся в них длительное время, на уровне сообщества отсутствует дефицит минеральных веществ, даже если интенсивность роста каждого индивидуума сама по себе почти всегда лимитируется питательными веществами, что имеет значение и для продукции биомассы на единицу площади. Упомянутая в разделе 6.2.2.2 специфичная для каждого элемента симптоматика, представляющая собой кратковременную реакцию на дефицит элемента питания, весьма существенна для сельскохозяйственных растений. При конкуренции видов и генотипов в местообитании остаются, однако, на долгое время, как правило, лишь те таксоны, которые могут справиться с ситуацией дефицита так, что подобных симптомов у них не проявляется. Удивительно, что с помощью анализа на элементы у дикорастущих растений можно лишь в редких случаях выяснить, есть ли дефицит минеральных веществ и, если есть, какого вещества именно. Эти растения растут таким обра ¦ зом, что дело не доходит до дефицита существенных питательных веществ, поскольку при известных условиях их органы, образующиеся в меньшем количестве и меньших размеров, полностью функциональны. В таких случаях рост соответствует наличию ресурсов. Рост, превышающий ресурсные возможности, быстро бы элиминировал такой вид или генотип благодаря снижению жизненности. Растения очень холодных биомов (высокогорья, полярные области), у которых поступление азота затруднено, имеют даже более высокие концентрации азота в листьях, чем сравнимые таксоны из теплых стран, что Т.Чепин назвал «растительным потреблением» (от англ. luxurious consumption). Условия этих жизненных пространств не дают возможности выживать с плохо снабжаемыми, малопродуктивными листьями. У таких приспособившихся видов оптимальное (экономное) потребление ограниченных ресурсов обеспечивается контролируемым ростом.
Еще по теме Стратегии инвестиций азота:
- ВЗАИМОДЕЙСТВИЕ АЗОТА, ФОСФОРА И КАЛИЯ В ПИТАНИИ РАСТЕНИЙ ПРИ ИСПОЛЬЗОВАНИИ ИМИ НИТРАТНЫХ И АММОНИЙНЫХ ФОРМ АЗОТА [22]
- Стратегии жизненных циклов
- R-K-стратеги и ценофобные-ценофильные виды
- 3. 2. Типы экологических стратегий
- Эколого-ценотические стратегии вилов и популяций растений
- Классификации, рассматривающие три жизненные стратегии
- Стратегия фуражировки
- ТЕХНОЛОГИЧЕСКАЯ СТРАТЕГИЯ
- Моя стратегия защиты
- 1.2. СТРАТЕГИЯ ЖИЗНИ. ПРИСПОСОБЛЕНИЕ, ПРОГРЕСС, ЭНЕРГЕТИЧЕСКОЕ И ИНФОРМАЦИОННОЕ ОБЕСПЕЧЕНИЕ
- ГЛАВА 8 Стратегии для городов и деревень
- КРУГОВОРОТ АЗОТА
- ИММОБИЛИЗАЦИЯ АЗОТА
- Источники и накопители азота
- КРУГОВОРОТ АЗОТА