ИЗУЧЕНИЕ ВНУТРИСЕМЕЙНОГО ПЕРИОДА ЭМБРИОГЕНЕЗА
Сравнительное изучение внутрисемейных стадий эмбриогенеза при знании степени и формы редукции позволяет ответить на вопрос о способах эволюционных морфогенетических преобразований.
Сем. Scrophulariaceae. Единство способа эмбриогенетического развития подтверждено для всех исследованных автотрофных и паразитных видов (Soueges, 1921, 1935с; Jyengar, 1940а, 1940b, 1940с, 1942а, 1942b,
1947; Raghavan, Srinivasan, 1941a, 1941b; Crete, 1948, 1950a, 1950b, 1950c, 1953, 1954a, 1954b, 1955, 1958; Berg, 1954; Tiagi, 1956; Arekal, 1963a, 1963b, 1963c, 1964; Никитичева, 1966, 1970; Никитичева, Терёхин, 1973, и др.)* Рис. 14 и 15 иллюстрируют начальные этапы эмбриогенеза у автотрофных и паразитных представителей семейства. При обсуждении данных о начальных этапах эмбриогенеза в сем. Scrophulariaceae необходимо обратить прежде всего внимание на способ дифференциации гипофиза у этих растений.
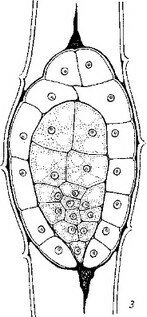
3. И. Никитичева (1966, 1970), предприняв исследование эмбриогенеза Норичниковых (Lathraea squamaria и некоторых других видов), показала, что представление Джогансена (Johansen, 1950) и ряда других исследователей о том, что гипофиз у Норичниковых дифференцируется обычно посредством образования двух специфических клеток («линзовидной» и «часового стекла»), основано на недостаточно детальном анализе первых этапов эмбриогенеза. В действительности же способ образования гипофиза посредством заложения двух косых, взаимно перпендикулярных перегородок, который рассматривался Джогансеном как крайне редкое исключение из общего правила, должен быть признан единственным способом образования гипофиза у Норичниковых. Наши исследования эмбриогенеза Bhynehocorys orientalis, Tozzia alpina и Melampyrum nemorosum полностью подтвердили представления, развиваемые 3. И. Никитичевой (рис. 16). Как видно из данных рис. 14, 15 и 16, первые этапы эмбриогенеза и способ дифференциации гипофиза едины у всех Норичниковых.
Дифференциация гипофиза происходит всегда таким образом, как это было показано 3. И. Никитичевой.Рассмотрим подробно эмбриогенез автотрофного растения Verbaseum thapsus (рис. 14). Среди основных особенностей эмбриогенеза этого растения следует упомянуть такие его черты, как характерное поперечное деление зиготы, Т-образную тетраду бластомеров, а также весьма обычные для покрытосеменных типичные стадии квадрантов и октантов. Дифференциация гипофиза у V. thapsus происходит способом, описанным ранее И. Никитичевой (1966) для Lathraeasquamaria1 т. е. посредством последовательного заложения в инициальной клетке гипофиза двух косых, взаимно перпендикулярных перегородок. Верхняя (линзовидная) клетка делится затем снова вертикальными перегородками на четыре доли, формируя таким образом «iec» — инициали периблемы, а клетка «часового стекла» дифференцируется в инициали чехлика корня.
Дифференциация гипофиза происходит примерно в тот же период, что и дифференциация эмбриодермы в клетках октантов. В период завершения дифференциации периблемы и плеромы начинается регионализация семядолей, т. е. возникновение морфологически выраженных примордиев семядолей. Это стадия «сердечка». На стадии «торпеды» дифференцируется зачаток стеблевого апекса (эпикотиль), который в зрелом семени морфологически выражен у зародыша в виде полусферы, лишенной зачатков первых листьев.
Эмбриогенез других автотрофных, а также и паразитных представителей сем. Норичниковых в своих основных чертах соответствует эмбриогенезу, описанному у V. thapsus. У изученных нами Rhynchoeorys orientalis, Melampyrum nemorosum и Tozzia alpina, как и у F. thapsus, образуется Т-образная тетрада бластомеров, стадии квадрантов и октантов, дерматоген и гипофиз, периблема и плерома (рис. 16).
Арекал (Arekal, 1963а) описал несколько иной способ эмбриогенети- ческого развития у Melampyrum Iineare. Он придал этому большую филогенетическую значимость. Однако то обстоятельство, что на первых этапах развития, в период дифференциации гипофиза и далее (как об этом можно судить по рисункам автора), строение и способ развития зародыша М.
Iineare в обшем соответствуют таковому у других Норичниковых (рис. 15), свидетельствует против выводов Арекала.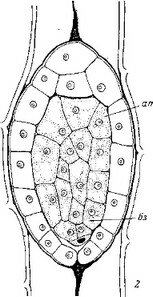
Рис. 14. Начальны© и средние стадии эмбриогенеза некоторых автотрофных представителей сем. Scrophulariaceae.
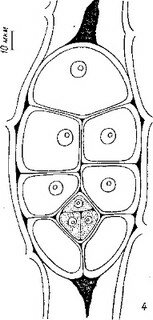
Рис. 14 (продолжение).
Э. С. Терёхин
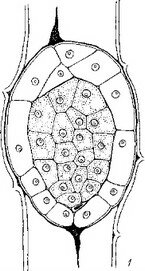
Рис. 15. Начальные и средние стадии эмбриогенеза некоторых паразитных представителей сем. Scrophulariaceae.
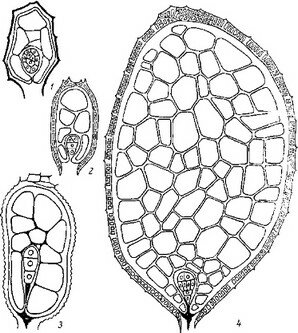
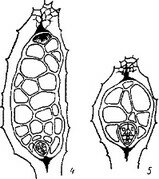
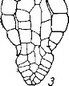
 I—25 — последовательность стадий, са — апикальная клетка проэмбрио; св — базальная клетка проэмбрио; инц — инициали центрального цилиндра; инч — инициали корневого чехлика.
I—25 — последовательность стадий, са — апикальная клетка проэмбрио; св — базальная клетка проэмбрио; инц — инициали центрального цилиндра; инч — инициали корневого чехлика.

Рис. 17. Начальные и средние стадии эмбриогенеза некоторых представителей
сем. Orobanchaceae
|
Вид |
Стадия диады |
Стадия тетра ды |
Стадия кбад- рантоВ |
Стадия октан- тод |
Образобание гипофиза |
||||
|
Aphyllon unif Iorum (Orobanche unifIora) (по Smithf 190k) |
§ |
(о|о) Ywl |
/6ТЭ\ 1оДо) |
||||||
|
Aphyllon unif I arum (Orobanche unif lord) (по Casserа, 1935) |
_г) ^ I чу/ |
/оТо\ щ |
|||||||
|
Orobanche pallidiflora (по НикитичеВой, 1970) |
Q о |
2 |
(сь) YoJ Yoi I M |
/§35\ KoV |
/|Rgt;\ f\ffy п I у и9) I 113(10) |
||||
|
Aegrineiia in die а (по Juliano, 1935) |
-OJ Oj I 72 |
I ~РI
1 I I 73 |
(о J юга Ygl П 74 |
[о о) (о Aj I 175 |
\ CD I W 78 |
||||
|
Aeg-inetia indica (по Tiagri, 1951) |
[оо] Го I |
(5) Io [о/ Yol оI |
I О } I00) Wj Oj \ PD I |
Гао\ .' GD^ [о I ¦Qj Q j ^ |lt;§у/ |
о\ lt;?/ W |
||||
У сравнительно слабоспециализированных паразитных видов, таких как Euphrasia arctica (Arekal, 1963а), Tozzia alpina, Rhynchocorys orientalis и других, сходство эмбриогенезов между собой и с непаразитными Норичниковыми может быть прослежено на всех стадиях развития зародыша — от зиготы до зрелого семени (рис. 15, 16). У названных растений сформированные зародыши имеют все основные органы и ткани, которые характерны для зародышей автотрофных Норичниковых: семядоли с васкулярным пучком в них; эпикотиль, иногда с примордиями первых листьев, как у Rhinanthus или Tozzia; гипокотиль с отчетливо выраженным васкулярным пучком, радикулу с чехликом и инициалями периблемы и плеромы; ткань эмбриодермы.
Таким образом, эмбриогенез слабоспециализированных паразитных Норичниковых во всех особенностях соответствует эмбриогенезу автотрофных представителей семейства.
Сравнение эмбриогенеза автотрофных и слабоспециализированных паразитных Норичниковых с эмбриогенезом переходных и высокоспециализированных паразитных растений из того же семейства, таких как Striga orobanchoides (Tiagi, 1956) и Lathraeasquamaria (Никитичева, 1966, 1970), показывает, что при сходстве эмбрионального развития на первых этапах зародыши слабоспециализированных и высокоспециализированных Норичниковых различаются уровнями дифференциации (по состоянию в зрелом семени).
Отсюда следует единственно возможный вывод, что редукционные изменения зародышей Норичниковых происходят путем элиминации конечных этапов развития той или иной эмбриональной структуры. Следуя
терминологии А. Н. Северцова (1949), мы можем сказать, что редукция структур зародышей происходит путем последовательной терминальной аббревиации. Эта последовательность, постепенность редукции отчетливо видна в строении зародышей в семенах растений-паразитов с разной степенью специализации (см. рис. 3 и 4).
Сем. Orobanchaceae. Изученные представители семейства характеризуются сходством способа эмбриогенеза не только между собой (рис.
14, 18), но и с представителями сем. Норичниковых (Cassera, 1935; Tiagi, 1950, 1951b, 1952а, 1952Ь, 1956, 1963; Crete, 1955; Терёхин, 1966, 1973; Ники- тичева, 1970, и др.)., Эмбриогенез, строение и степень развития зрелых зародышей у Cistanche tubulosa и у Striga orobanehoides в общем идентичны. В связи с этим мы можем считать, что оба эти растения находятся примерно на одном уровне эволюционных преобразований. Однако сравнение эмбриогенезов S. ого- banchoides и С. tubulosa с эмбриогенезом других заразих (Phelypaea cocci- пеа, Orobance colorata, О. purpurea и Phacellanthus tubiflorus) показывает, что при общности начальных и промежуточных этапов эмбриогенеза зародыши Phelypaea, Orobanche, Phacellanthus имеют меньшую дифферен- цированность в зрелом состоянии (не всегда полную дифференциацию периблемы и плеромы, меньшее число клеток в этих тканях и в эмбриодерме). Очевидно, здесь имеет место следующая ступень эволюционного недоразвития, вызванного элиминацией завершающих стадий развития. Модус последовательной терминальной аббревиации отчетливо проявляется и на этом этапе эволюционных преобразований.
У последнего звена эволюционного ряда Заразиховых, у Aeginetia indica, мы обнаруживаем крайнюю степень редукции зародыша (рис. 6, 6). Джулиано (Juliano, 1935) и Тиаги (Tiagi, 1952b) отметили в отношении Aeginetia, что после стадии Т-образной тетрады бластомеров здесь нельзя обнаружить характерных фигур квадрантов и октантов, не образуется гипофиз, не дифференцируется периблема и плерома. Вместо этого у Aegi- netia можно видеть более простые вариации дробления, прямым путем приводящие к становлению структуры глобулярного зародыша, образованного из меньшего, чем у предыдущих видов, числа клеток, при весьма слабой дифференциации эмбриодермы (рис. 17).
Тиаги (Tiagi, 1952b) на основании того, что суспензор Aeginetia также имеет редуцированное развитие и состоит всего лишь из одной клетки, счел возможным трактовать эмбриогенез Aeginetia в отличие от эмбриогенеза остальных Заразиховых, как эмбриогенез типа Caryophyllad.
Однако вся совокупность признаков эмбрионального развития Aeginetia свидетельствует против такого заключения, особенно в связи с значительной вариацией числа дериватов от сЪ, обнаруженных нами и у других заразих (Терёхин, 1966).Весьма существенно и то обстоятельство, что эмбриогенез Aeginetia и на средних этапах весьма вариабелен. Однако ослабление жесткой детерминированности эмбриогенеза, связанное, очевидно, с утратой строгой гистологической организации в период созревания семени, можно наблюдать не только у Aeginetia, но и у других Заразиховых (правда, на более поздних этапах эмбриогенеза). У Заразиховых это явление обычно выражается в нарушении правильного дробления этажа I при дифференциации эмбриодермы. Создается впечатление, что в эмбриогенезе Заразиховых дифференциация этажа I (в противоположность таковой у Норичниковых) не подчинена сколько-нибудь жесткому контролю со стороны генотипа и совершается с высокой степенью вариабельности. У Aegi- netia явление ослабленной детерминированности распространяется на более ранние этапы эмбриогенеза и проявляется уже на стадии квадрантов.
Можно думать, что состояние ослабленной детерминированности всегда предшествует процессам редукции. Г. А. Левитский (1924—1925, стр. 116), изучив редукцию половых органов у спаржи, объяснил значи-
Рис. 18. Эмбриогенез PMypaea coccinea (сем. Orobanchaceae)
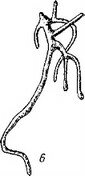 - последовательность стадий, а - энное™™- я ™спасеае).
- последовательность стадий, а - энное™™- я ™спасеае).
эндосперм, б - зародыши; . _ семенная оболочка.
тельное варьирование развития рудиментарного пестика утратой последним своего функционального значения. Вследствие этого пестик вышел из-под контроля естественного отбора, поддерживающего должную детерминированность лишь у функционирующих органов.
Из приведенных выше примеров явствует, что структурная редукция зародышей Заразиховых так же, как и редукция зародышей у Норичниковых, происходит путем последовательных этапов терминальной аббревиации. Нельзя не отметить при этом, что логичным следствием такого типа процессов на первых этапах эволюции являются преобразования посредством общей редукции биомассы зародышей (Rhynchocorys, Tozziay Striga Ъаитапи). В дальнейшем в зависимости от характера изменений в функциональной деятельности процессы элиминации происходят более активно в одних структурах и менее активно в других (дифференциальная редукция).
Сем. Balanophoraceae. Среди растений группы эктопаразитов наиболее редуцированные зародыши обнаружены у видов родов Balanophora и Langsdorffia. Детальное изучение эмбриогенеза у Balanophora globosa и некоторых других видов этого же рода (Терёхин, Яковлев, 1967) позволило выявить одну интересную закономерность, на которую не обратили внимания предшествующие исследователи (Hofmeister, 1858; Treub, 1898; Lotsy, 1899; Ernst, 1914; Ekambaram, Panje, 1935; Zweifel, 1939). Скованные сложившимися представлениями, названные исследователи дискутировали вопрос о том, продольно или поперечно делится зигота у Balano- phora. Нам удалось выяснить, что в споре были правы обе стороны, поскольку деление зиготы Balanophora правильнее отнести к неориентированному способу дробления (рис. 19). Это означает, что деление в зиготе Balanophora может происходить в любой ориентации по отношению к самой зиготе или ее положению в зародышевом мешке (эндосперме).
Следующий этап эмбриогенеза Balanophora — образование посредством двух взаимно перпендикулярных делений характерной стадии квадрантов. Затем следует дифференциация эмбриодермы. На этом эмбриогенез собственно заканчивается, или же некоторые из вновь образовавшихся клеток могут поделиться еще однажды (рис. 19). Результат эмбриогенеза — образование немногоклеточного овального тела. Отсутствие жесткой детерминации в способе дробления бластомеров, столь характерное для Orobanehe на последних стадиях эмбриогенеза, а для Aeginetia также и на его средних стадиях, у Balanophora проявляется уже при первом неориентированном (лабильном) делении зиготы. Пример Balanophora, таким образом, еще раз подтверждает общую закономерность в изменениях эмбриогенеза паразитных покрытосеменных, описанную нами выше.
Сем. Loranthaeeae9 подсем. Viseoideae. Способ дробления зиготы (поперечное деление), ранний эмбриогенез и основные черты строения зрелых зародышей принципиально идентичны у представителей обоих изученных родов — Viseum и Areeuthobium (Pisek, 1923; Steindl, 1935; Cohen, 1963). Строение базальной области зародыша у обоих родов растений весьма характерно и очень сходно: масса меристематических клеток окружена сосочковидным эпидермисом. Однако при сравнении зародышей Viseum album и Arceuthobium, oxyeedri разительно выступают их различия в степени дифференцированности апикальной области.
Учитывая принципиальное сходство этих растений в способах эмбриогенеза, нельзя не прийти к заключению, что эволюционные изменения зародыша происходили и здесь посредством терминальной аббревиации.
Еще по теме ИЗУЧЕНИЕ ВНУТРИСЕМЕЙНОГО ПЕРИОДА ЭМБРИОГЕНЕЗА:
- ИЗУЧЕНИЕ ВНУТРИСЕМЕЙНОГО периода РАЗВИТИЯ ЗАРОДЫША
- ВНУТРИСЕМЕЙНОЙ ПЕРИОД РАЗВИТИЯ ЗАРОДЫША
- §2. Периоды онтогенеза 2.1. Пренатальный
- ПОСЛЕРОДОВОЙ ПЕРИОД
- ПОСЛЕРОДОВОЙ ПЕРИОД
- Осенний период.
- Летний период.
- 7.1. ЭТАПЫ. ПЕРИОДЫ И СТАДИИ ОНТОГЕНЕЗА
- 9.1. КРИТИЧЕСКИЕ ПЕРИОДЫ В ОНТОГЕНЕЗЕ ЧЕЛОВЕКА
- Период греко-римский
- Ранне - весенний период .
- Период рождения молодняка.
- ПАТОЛОГИИ ПОСЛЕРОДОВОГО ПЕРИОДА
- ПРОФИЛАКТИКА БОЛЕЗНЕЙ В ПАСТБИЩНЫЙ ПЕРИОД
- Поздне - весенний период.
- Новый период.
- ГЛАВА IV НАЧАЛЬНЫЙ ПЕРИОД РАЗВИТИЯ ЖИЗНИ
- Развитие экологии в последарвиновский период.
- ДОБИОСФЕРНЫЙ ПЕРИОД ПЛАНЕТЫ ЗЕМЛЯ
- Период средних веков.