Вертикальная зональность донной растительности
Для морской донной растительности характерна вертикальная зональность распределения как отдельных видов, так и растительных группировок. Исследованию закономерностей этой зональности, её связи с биотическими и абиотическими факторами посвящены сотни работ, выполненных в самых разных частях Мирового океана, в них предложены несколько схем вертикальной зональности.
Наибольшее распространение получили схемы, основанные на учёте уровней приливов и отливов и других характеристик абиотической среды, в первую очередь прибойности. Например, верхняя литораль ограничена уровнями сизигийного и квадратурного приливов, средняя — уровнями квадратурных прилива и отлива нижняя — уровнями квадратурного и сизигийного отливов. Очевидно, что главным фактором в распределении жизни здесь выступает время увлажнения того или иного участка дна. Этот способ достаточно хорошо работает лишь в регионах с выраженными приливами и отливами (Перестенко, 1980).Первую схему, основанную на характеристиках абиотической среды, разработал L. Vaillant (1891, цит. по: Возжинская, 1977). Наиболее распространена схема F.R. Kjel- lman (1877). Исследуя флору Мурмана, он выделил литораль, сублитораль и элитораль (он же, кстати, первым ввёл и термин «формация» для обозначения группировок растительности). Элиторалью он назвал зону, расположенную ниже максимальных глубин распространения растительности. Ещё раньше J.R. Lorenz (1863) ввёл в обиход термин «супра- литораль» (обе ссылки по: den Hartog, 1959). Эти термины гидроботаники используют до сих пор, что вызывает определённую путаницу, поскольку в работах зоологов под сублиторалью обычно понимают сублитораль и элитораль вместе. Разные исследователи растительности вносят свои коррективы и дополнения, связанные, в основном, с тем, работают ли они в приливных или бесприливных морях. Так, средиземноморская школа гидробиологов (в первую очередь, J.M.
Peres), имея дело с практически бесприливным морем, выделяет супралитораль (над уровнем моря), узкую эулитораль (около нуля глубин) и сублитораль, подразделяемую на горизонты или этажи: инфралитораль (от уровня моря и до 25-30 м) и циркалитораль — зону, где уже нет растительности (глубже 30 м). При высо-о
•lt;
S'
малая вода при квадратурных отливах
корковые водоросли и лишаиники
(Prasiola, Verrucaria)
нитчатые водоросли (Bangia, Urospora)
пластинчатые водоросли , (Porphyra, Enteromorpha) Pelvetia
супралитораль
полная вода при_ сизигийных приливах
полная вода при квадратурных приливах
малая, вода. при_
сизигийных отливах
Laminaria digitata
Laminaria hyperborea
кустистые багрянки
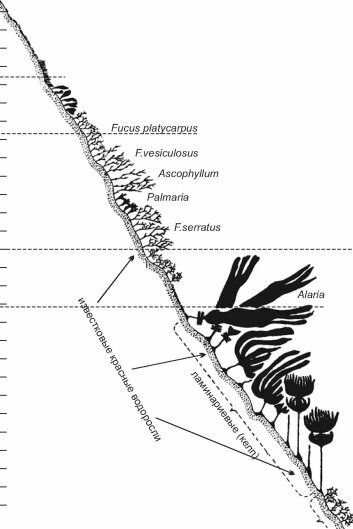
Рис. 3.28. Схема распределения водорослей на скалистом берегу Па-де-Кале. По Gessner (1955).
+14 +13 +12 +11 +10 +9 +8 +7 +6 +5 +4 +3 +2 + 1 0 -1 -2 -3 -4 -5 -6 -7 -8 -9 -10 -11 -12 -13 -14 -15
кой прозрачности средиземноморских вод макрофитобентос часто распространяется до 100 м и более, и верхняя граница циркалиторали смещается на соответствующую глубину (Ercegovic, 1980; Cebrian, Ballesteros, 2003 и многие другие).
Супралитораль (зона заплеска) расположена над уровнем моря или выше линии максимального прилива. Её увлажняют только набегающие волны, и очень редко (во время нагонных ветров) она совсем скрывается под водой. В некоторых местах с сильным волнением (например, на Фаррерских о-вах) она достигает 25 м ширины. Супралитораль населяют немногочисленные водоросли, образующие корки, подушки и густые дерновины, позволяющие сохранять влагу, а также некоторые лишайники. В Белом море в этой зоне обитает фукоид Pelvetia canaliculata; в Чёрном, в зимний период, когда шторма особенно часты и сильны, здесь развивается «зимняя» растительность, представленная примитивными багрянками Nemalion и Bcmgia.
Литораль иногда эту зону именуют эулитораль (т. е. собственно литораль, настоящая литораль). В верхней литорали обитают растения, способные переносить долгое осушение. Это многочисленные галофиты, синезелёные водоросли, зелёные и бурые нитчатки, ряд багрянок с корковыми или сильно ослизнёнными талломами. В Белом море сюда поднимаются разреженные заросли Fucus vesiculosus, который способен переживать резкие перепады температуры и иссушение вплоть до воздушно-сухого состояния. В средней литорали периоды затопления и обсыхания практически одинаковы, и флора здесь существенно богаче как качественно, так и количественно. В умеренных и бореальных водах здесь развиваются мощные заросли F. vesiculosus с вкраплением куртин Ascophyllum nodosum (проективное покрытие достигает 60-80%) с многочисленными эпифитами и «подлеском» из багрянок и зелёных водорослей. Нижняя литораль ещё богаче жизнью, доминирующие здесь фукоиды крупнее, их флора богаче (добавляются F. serrcitus, F. dis- tichus), проективное покрытие дна достигает 100%. Отметим, что для северных морей характерна ярко выраженная поясность распределения растительного покрова, которая чётко прослеживается уже в осушной зоне.
В Чёрном море (а также Японском, Средиземном и некоторых других) литораль не выражена, лишь иногда при сгонных ветрах обнажается узкая полоска дна, покрытая мозаикой низкорослых видов (багрянки Ceramium ciliatum, Polysiphonia ораса, бурые Padina pavonia, Dilofus fasciola, проростки цистозиры, зелёная Enteromorpha spp.). Эту зону называют псевдолиторалью. В Средиземном море она занята в основном багрянками (Нур- пеа musciformis и др.), имеющими совершенно не характерную для красных водорослей окраску: ярко- или сине-зелёную.
Отечественные исследователи (К.М. Дерюгин, Е.Ф. Гурьянова, П.В. Ушаков, Т.Ф. Щапова, О.Б. Мокиевский, Л.П. Перестенко, О.Г. Кусакин), работавшие на Мурманском побережье и на Дальнем Востоке, выделяют несколько биономических типов литорали в зависимости от степени прибойности: I — полузащищённый берег, II — открытый морской берег, III — открытый океанский берег, IV — лагунный, V — эстуарный и VI — литоральных ванн (Перестенко, 1980; Kussakin, 1977).
Для каждого из них характерна определённая зональность фито- и зообентоса, причём в разных морях конкретные пояса биоты часто образованы сходными викарирующими видами или группировками видов. Так, в нижней литорали открытого побережья Мурмана доминируют ламинариевые Alaria esculenta и Laminaria digitata, а в аналогичных по биономическому типу местообитаниях Камчатки и Северных Курил —Alaria angusta и Laminaria longipes.Сублитораль подразделяют на два горизонта по степени освещённости — горизонт фотофильной растительности (от 0,1 м до примерно 15-25 м глубины) и горизонт сциа- фильной растительности (от примерно 25-30 м и до нижней границы фитали). Фото- фильный горизонт — «царство» морских трав и самых крупных водорослей — бурых ламинариевых с широкими и длинными пластинчатыми талломами (Laminaria, Alciria, Aga- rum, Macrocystis, Lessonia, Nereocystis и др.), которым сопутствуют многочисленные эпифиты и растения «подлеска». В тёплых и тропических морях на этом горизонте ламинариевых сменяют фукусовые (Sargassum, Cystoseira, Turbinaria, Hormosira и др.). Сциа- фильный горизонт обжит теневыносливыми красными водорослями, ярким примером которых служит черноморская Phyllophora crispa (= Ph. nervosa), особенно её пластообразующая форма, встречавшаяся на Филлофорном поле Зёрнова до глубины 90 м. Горизонты, в свою очередь, подразделяют на этажи.
Существуют также классификации, основанные не на абиотических факторах, а на распределении поясов биоты. Одну из таких классификаций разработали супруги Т.А. и Т. Stephenson (1949; цит. по: den Hartog, 1959) на основе своих более чем 30-летних исследований прибрежной биоты в Великобритании, Южной Африке, Красном море, на Большом Барьерном Рифе и на атлантическом и тихоокеанском побережьях Северной Америки. При всём разнообразии этих местообитаний удалось выделить общие характерные черты вертикального распределения донных растений и животных. Стефенсоны выделили следующие зоны: супралиторальную кайму (supralittoral fringe), литораль (midlittoral zone), инфралиторальную кайму (infralittoral fringe) и собственно инфралиторальную зону, простирающуюся до границы фитали или всего шельфа.
Границы между этими зонами определены по массовым видам биоты: балянусам, литторинам, крупным водорослям. Схема Стефенсонов нашла как множество последователей, так и критиков, поскольку её применение более трудоёмко.Говоря о вертикальной зональности донных растительных сообществ следует учитывать особенности их распределения на разных элементах подводного рельефа. Так, доминирующей формой рельефа дна у побережья Северного Кавказа (Чёрное море) является грядовый бенч: чередование поднятий — гребней, сложенных в данном случае осадочными породами (мергелем), и депрессий — лощин или расщелин. П.В. Рыбников (1993а, 1997) показал, что характер сообщества обрастания багрянки Phyllophora nervosa существенно различается на гребнях и в лощинах. При этом микрорельеф дна играет более значительную роль, чем глубина, и сообщества гребней более «мелководные» по составу и структуре, а сообщества лощин — более «глубоководные». Фауна нижних горизонтов фитали проникает в более мелководные участки по лощинам, а мелководная фауна продвигается вглубь по гребням. Распределение эпифитной флоры также зависит от микрорельефа дна: в защищённых лощинах она богаче, и экологически лощины более «глубоководны», чем гребни на той же глубине (Лучина, Рыбников, 1993). Что касается самой филлофоры, то на гребнях у неё образуется больше молодых сегментов, и их доля в общей массе растения выше. Так, на гребнях на глубине 1 м прирост последнего года составляет в среднем 13% массы растения, а в расщелинах на той же глубине — всего 6%. Удельное число молодых сегментов также выше на гребнях, чем в лощинах: на глубине 1 м 140 шт/г и 60 шт/г, соответственно. С глубиной (от 1 до 10 м) этот показатель убывает для обоих элементов, но в разном диапазоне значений: на гребнях — от 140 до 60 шт/г, в лощинах — от 60 до 30 шт/г (Рыбников, Лучина, 1998). Основным фактором, обеспечивающим эти различия, авторы считают интегральную подвижность воды, гораздо более интенсивную на гребнях, чем в защищённых гребнями лощинах. Кроме того, после штормов в лощинах надолго остаётся холодная вода, занесённая из-под термоклина, а насыщение воды кислородом на гребнях выше, чем в расщелинах, что не может не сказываться на состоянии сообществ, особенно зооэпифитона (Рыбников, 19936).
Таким образом, характеризуя донные сообщества нельзя ограничиваться определением одной лишь глубины. Пробы, отобранные с одинаковой глубины, но с разных элементов рельефа, могут различаться так же сильно, как пробы с разных глубин. Игнорирование таких особенностей местности может привести к серьёзным ошибкам, например, при оценке фонового состояния биоты перед строительством хозяйственных объектов. Впоследствии, на этапе экспертизы воздействия строительства на окружающую среду, эти изначальные ошибки повлекут за собой неверную оценку нанесённого ущерба.
Еще по теме Вертикальная зональность донной растительности:
- Вертикальная зональность
- Ботанико-географическая зональность
- Общие закономерности вертикальной стратификации микробных сообществ
- БИОЦЕНОЗЫ ЗОНАЛЬНЫХ ТИПОВ ПОЧВ СССР
- Вертикальная структура
- Вертикальные миграции рыб в морских водоемах
- Зонально-подзональные варианты строения торфяных отложений
- Зональные особенности гумусообразования в подзолистых почвах Кольского полуострова
- Динамика вертикальной скорости торфонакопления, биологической продуктивности болотных систем и темпов заболачивания
- НАУЧНЫЕ ПРИНЦИПЫ ЗОНАЛЬНЫХ СИСТЕМ ПРИМЕНЕНИЯ УДОБРЕНИЙ В СЕВООБОРОТАХ
- Образование льда Вертикальная циркуляция и образование льда в пресной воде
- Классификация морских донных растительных сообществ
- Растительность