Сукцессии и взаимоотношения макрофитов
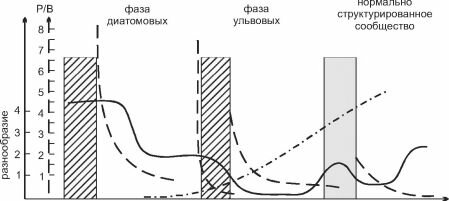
При освоении нового субстрата растительность проходит несколько сукцессионных стадий (рис. 3.44). Первыми (в течение суток) твёрдый субстрат заселяют диатомеи, причём как бентосные, так и планктонные.
Последние, однако, через неделю исчезают из сообщества. Для этой стадии характерно очень высокое видовое разнообразие. Через 9-12 дней появляются первые макрофиты. Все они — высокопродуктивные быстрорастущие оппортунисты с нитчатыми или трубчатыми талломами: зелёные Ulothrix, Enteromorpha spp., красные Ceramium spp., Polysiphonia и др. Развиваются и вертикальные колонии ди- атомей (Navicula). Эта стадия сукцессии длится 30-120 дней от начала заселения, её называют фазой ульвовых, поскольку преобладающими в ней являются ульвовые зелёные водоросли. В переходном периоде между первой и второй фазами видовое разнообразие диатомей падает, их покров из сплошного становится пятнистым, и постепенно первые «колонизаторы» исчезают. Третья фаза (нормально структурированного сообщества) характеризуется появлением и развитием многолетних форм с меньшими скоростями роста (она начинается примерно через 3 месяца). Изменения во флористическом составе сообщества сопровождают существенные функциональные изменения при развитии сукцессии: видовое разнообразие падает к концу каждой фазы и возрастает в начале следующей при общем тренде увеличения; коэффициент P/В стремительно падает от первой фазы к третьей; концентрация хлорофилла а, напротив, существенно возрастает. Показано, что в переходные моменты от одной фазы к другой увеличивается поток энергии через сообщество, а рассеяние энергии последовательно уменьшается от первой фазы к третьей. При этом растительность третьей фазы зависит от степени развития растительности второй, в то время как фаза ульвовых не зависит от первой фазы, и интенсивность её развития определяется только стратегиями роста её компонентов (Niel, Valera, 1984). Таким образом, взаимовлияние макрофитов начинает сказываться на ранних стадиях сукцессии.сукцессионная смена происходит с перекрыванием сообществ
Рис. 3.44. Сукцессия водорослей на новом субстрате. По Niell, Valera (1984).
СтрУктУРУ фитоценозов определяют биотические взаимодействия и абиотические факторы. Биотические взаимодействия могут как быть в пределах фитоценоза (конкурен-

преимущество в потреблении
химическое
подавление
нарастание
биогены
перекрывание доступа к субстрату
споры Ulothrix
Pseudolithophyllum
гипотетический случаи
Lithothamnion
Gracilaria Enteromorpha дерновина Enteromorpha
Рис. 3.45. Механизмы реализации конкуренции. По Olson, Lubchenco (1990).
ция за субстрат, свет, биогены и т. д.), так и растениями и остальными компонентами сообщества.
Конкуренция влияет на возрастную и размерную структуру и успех размножения отдельных особей. Конкурентоспособность макроводорослей определяется всем комплексом их характерных черт, как морфологических (форма и степень разветвлённости таллома, его толщина и прочность), так и физиологических (чувствительность к свету, способы поглощения и запасания питательных веществ, выделение защитных химических соединений, способность к модификациям) (Olson, Lubchenco, 1990).
Можно выделить несколько механизмов реализации конкурентных способностей (рис. 3.45): способность к интенсивному потреблению питательных веществ; способность к быстрому захвату субстрата с образованием плотных зарослей-дерновин (turf); способность к непосредственной топической конкуренции — т. е. к механическому вытеснению конкурентов через нарастание на их талломы; аллелохимическое воздействие на конкурентов (через антибиотики и токсины).
Первый механизм иллюстрируют взаимоотношения зелёной ульвовой Enteromorpha sp. и красной Gracilaria tikvahiae. Если содержание питательных веществ в среде постоянно, то r-стратег энтероморфа вытесняет грацилярию за счёт существенно более высокой скорости потребления. Но грацилярия имеет более высокую запасающую способность, поэтому при значительных колебаниях концентрации питательных веществ она получает конкурентное преимущество и вытесняет энтероморфу.
Второй механизм свойственен дерновино-образующим водорослям. На рисунке представлена ситуация, при которой споры Ulothrix просто не имеют возможности достичь субстрата, сплошь покрытого дерновиной энтероморфы. Аналогичная картина наблюдается во взаимоотношениях энтероморфы с зиготами Fiiciis vesiculosus. Однако при появлении фитофагов ситуация меняется: фитофаги выедают пятна в покрове энтероморфы, зиготы фукуса оседают в них, и в течение некоторого времени фукус вытесняет энтероморфу, которая полностью исчезает под его покровом, но может перейти к эпифитиро- ванию на выросших талломах фукуса. Если же удалить фитофагов после оседания зигот фукуса, то энтероморфа довольно быстро «забьёт» проростки фукуса, закрыв их от света. Т. е. при определении конкурентных отношений между видами следует учитывать различную конкурентную способность разных онтогенетических стадий.
Третий механизм представлен нарастанием таллома одной корковой багрянки поверх таллома другой. Такое прямое уничтожение топического конкурента среди макроводорослей наблюдают редко. Однако примерно тот же механизм обеспечивает успешность инвазии описанных выше Caulerpa taxifolia и Sargassum muticum.
Наконец, четвёртый механизм подразумевает химическое подавление конкурента за счёт экзометаболитов. На рисунке авторы представляют его как гипотетический, однако в природе наблюдаются вполне реальные примеры такого воздействия. Талломы многих водорослей, особенно их молодые активно растущие части, выделяют антибиотики, препятствующие развитию эпифитов — конкурентов за свет и питательные вещества.
Выше упомянуто угнетающее воздействие экзометаболитов Caulerpa taxifolia на другие макроводоросли. Красные кораллиновые водоросли выделяют соединения, подавляющие рост и даже убивающие гаметофиты и ювенильные спорофиты ламинариевых (Denboh et al., 1997). В Японии обширные области в прибрежье Хоккайдо после массового развития морских ежей Strongylocentrotus nudus лишились коммерчески важных зарослей ламинариевых и теперь представляют собой пустоши, покрытые панцирем корковых и кустистых кораллиновых (Lithophyllum yessoense, Pneophyllum zostericolum, Corallina pilulifera). При достаточно высоких температурах (10-18°С) они выделяют броморганические соединения, из которых самым токсичным оказался бромоформ. Замещение ламинариевых кораллиновы- ми получило в Японии название «исояке» и вошло в международную гидроботаничекую терминологию. Восстановление зарослей ламинариевых происходит лишь при понижении температуры (до 6°С) во время спороношения и развития проростков, причём одновременно падает активность производства токсичных экзометаболитов у кораллиновых.Долгое время считали, что верхний предел вертикального распространения растительности определяют физические факторы (инсоляция, иссушение), а нижний — биологические взаимодействия. Однако оказалось, что биотические взаимодействия влияют и на положение верхней границы водорослевых поясов. Следует учесть, что как биотические взаимодействия, так и абиотические факторы имеют ещё и временную динамику — суточную, сезонную, межгодовую, долгопериодную. Необходимо также учитывать и особенности цикла развития самих макрофитов (Петров Ю.Е., 1974; Хайлов и др., 1992). Приведём несколько примеров.
В Великобритании удаление растительности выше поясов фукоидов {Fucus vesiculosus, F. serratus) и Laminaria digitata вызвало распространение этих видов вверх. Удаление растительности ниже поясов тех же водорослей, а также Pelvetia canaliculata, Himanthalia elongata, F. spiralis вызвало расширение поясов фукоидов вниз.
Следовательно, и верхняя, и нижняя границы поясов обусловлены в норме биотическими взаимоотношениями (Hawkins, Hartnoll, 1985).На скалистых побережьях Чили доминантом нижнелиторальной растительности является зелёный корковый Codium dimorphum. Его покров (проективное покрытие от 35% в январе до 62% в июле) располагается узкой полосой (50-100 см) выше пояса ламинариевых Lessonia nigrescens и Durvillea antarctica. Летом из-за высоких температур и повышенной солнечной радиации колонии кодиума обесцвечиваются и редуцируются вплоть до полной гибели. Освободившийся субстрат очень быстро колонизируют среднелиторальные водоросли (Ulva rigida, Iridaea boriana и др.), проникая, таким образом, глубже обычной нижней границы своего распространения (рис. 3.46). При искусственном удалении колоний кодиума наблюдается сукцессия, соответствующая приведённой в начале этого раздела схеме: первыми появляются диатомеи и цианобактерии, их быстро сменяет ульвовая фаза (16 видов верхнє- и среднелиторальных водорослей), а кодиум появляется в сообществе лишь несколько месяцев спустя. Если время его появления приходится на лето, то завоевать прежние позиции ему мешают фитофаги: они активно пожирают ослабленные обесцвеченные краевые участки колоний. Если же температурные условия не вызывают угнетения колоний кодиума, то он восстанавливает свой покров. Таким образом, на состояние пояса кодиума прежде всего влияют климатические факторы и фитофагия, а распространение вглубь более высоко растущих видов ограничивает только пояс кодиума (Santelices et al., 1981).
Codium dimorphum Регуляция нижней границы распространения во
1976 1977
Рис. 3.46. Динамика проективного покрытия на литорали Чили. По Santelices et al. (1981).
0
-8-
0
ct
другие водоросли
40-

80-
дорослей за счёт конкуренции была прослежена на скалистой литорали Новой Англии (США), где среднюю часть занимают заросли фукоидов (Fuchs vesiculosus, F distichus f.
edentatus), а в нижней части расположен пояс багрянки Chondrus crispus («ирландский мох»). Удаление хондруса привело к распространению фукоидов в нижнюю литораль, причём их состояние (размер, скорость роста, интенсивность размножения) на новом месте оказалось много лучше, чем в привычном биотопе. Следовательно, только конкуренция со стороны хондруса мешала фукоидам распространиться глубже. Наличие хондруса препятствует оседанию и прикреплению зигот фукусов как механически, так и за счёт экзометаболитов. Проростки фукоидов, а тем более взрослые растения индифферентны к соседству с хондрусом, т. е. конкуренция проявляется только на стадии пропагул.Интенсивность заселения освободившейся площади регулировали фитофаги (Littorina littorea): если при удалении хондруса их было мало, то фукоиды занимали до 100% освободившейся поверхности; если же моллюски были обильны, то образовывались разреженные заросли фукоидов. Таким образом, конкуренция определяет саму зональность литорали (наличие/отсутствие вида), а фитофагия — интенсивность развития вида внутри пояса. Что касается ограничения пояса Chondrus, то его верхняя граница определяется временем осушения, нормальным для фукоидов, но чрезмерным для багрянок, а нижняя — активностью морских ежей. Там, где ежей нет, хондрус способен проникать в сублитораль до глубины 10-20 м (Lubchenco, 1980).
На скалистом побережье Калифорнии на глубинах 0,3-2,3 м обитают ламинариевая Eisenia arborea, фукоид Halidrys dioica и багрянка Pterocladia capillacea (рис. 3.47). Крупные, до 80 см длиной, талломы эйсении образуют плотный полог (до 95% проективного
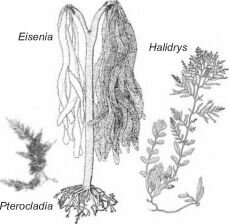
покрытия), под которым выживает птеро- кладия, способная жить в затенении под пологом эйсении (поглощающего до 90% поверхностной солнечной радиации). Для халидриса же этого количества света недостаточно. Фукоид занимает более высокий горизонт литорали, а также небольшие пятна внутри пояса эйсении, где она отсутствует, и его проективное покрытие здесь не превышает 20%. После удаления эйсении начинается бурный рост халидриса, который полностью вытесняет птерокладию за 6 месяцев. В тех местообитаниях, где хали- дрис редок или вовсе отсутствует, птеро- кладия (и ряд других водорослей из нижнего яруса зарослей эйсении) прекрасно раз- Рис. 3.47. Взаимодействующие вивается вне зарослей эйсении, но при на-
калифорнийские водоросли. По Abbott, личии халидриса она сосредоточена имен-
Hollenberg (1976). но ПОд спасительным пологом. Природное
Размеры водорослей пропорциональны природным.
разрушение зарослей эйсении сильными штормами привело к захвату освободившейся площади халидрисом, причём его проективное покрытие достигло 80%, а состояние зарослей выше и ниже разрушенного пояса эйсении не изменилось. Очевидно, что именно этот горизонт является оптимальным для халидриса. Таким образом, распространение Halidris определяет конкуренция с Eisenia, a Pterocladia развивается исключительно благодаря Eisenia (Kastendiek, 1982).
Еще по теме Сукцессии и взаимоотношения макрофитов:
- Сообщества морских макрофитов[‡‡‡‡‡]
- Макрофиты
- Макрофиты[††††]
- Макрофиты — вредные вселенцы
- Сукцессии в бентосе
- Деструктивные сукцессии
- Конструктивные сукцессии
- Дополнительные замечания О сукцессиях
- Общие закономерности сукцессий.
- МИКРОБНАЯ СУКЦЕССИЯ В ПОЧВЕ
- Васенев И.И., Таргульян В.О.. Ветровал и таёжное почвообразование. Режимы, процессы, морфогенез почвенных сукцессий, 1995
- СУКЦЕССИИ
- 5. Динамика биогеоценозов. Сукцессии
- Глава IV СУКЦЕССИЯ ВИДОВ МИКРООРГАНИЗМОВ ПРИ РАСПАДЕ ЕЛОВОГТ ДРЕВЕСИНЫ
- Продуктивность сообществ и жизненные стратегии водорослей. Морфофункциональный подход к изучению сообществ макрофитов
- Микрофлора сложных липняковых уборов. Сукцессия видов
- Пьявченко Н.И. (ред.). Взаимоотношения леса и болота, 1967
- Взаимоотношения животных между собой