Типы ареалов
Ареалы составляются путем объединения местонахождений одного таксона и лучше всего отражаются на картах ареалов (рис. 14.15. см. рис. 10.31, 11.259, 14.16 — 14.18).
Содержательность изображения ареала зависит от правильного систематического ограничения таксона или его отчле- нения от группы близкородственных видов, а также от достоверности флористических данных о его точках произрастания. Даже для ареалогического анализа флоры сосудистых растений Центральной Европы нет еще пока полной систематической и флористической основы.Картографическое представление о распространении таксонов нуждается в некотором абстрагировании, поскольку частота встречаемости и пространственное размещение индивидумов в мелких и более крупных местах обитания с учетом находящихся между ними промежутков обычно очень неравномерны Широко распространены точечные и контурные карты, или оба эти метода комбинируются (см. рис. 10.31, 14.25). При современных ареалогических исследованиях с применением электронных методов обработки данных (например, при проведении международных работ по картированию флоры Центральной Европы) особенно хорошо показали себя растровые карты, на которых наличие или отсутствие таксона в каждом случае наносилось на поле картирования определенного фиксированного размера (см. рис. 14.15). На высотных профилях можно также представлять вертикальное распространение определенного таксона.
Картирование флоры Центральной Европы Eryngium campestre L.
Ф Произрастание подтверждено + Вымер
Ф Вероятно вымер
? Случайный занос
q Статус под вопросом, старые сведения у Подтверждено недостаточно
0 _ 100
18

Рис. 14.15.
Пример растровой карты (предварительной) из проекта «Картирование флоры Центральной Европы»: синеголовник полевой (Eryngium campestre, Apiaceae) (по Н. Niklfeld).Представлено присутствие или отсутствие вида в полях градусной сетки длиной 10x6' географической долготы и соответственно шириной примерно 12x6 км. Субсредиземноморско-понтийский вид распространяется по теплым и сухим логам и долинам рек далеко на север
Для описания, сравнения и анализа ареалов таксонов важны следующие критерии:
- размер: от локального (эндемичного) до континентального и + распространенного по всему миру (космополитного);
- непрерывность: от полностью замкнутого (непрерывного) до сильно фрагментированного (дизъюнктивного);
- плотность заселенности: повсеместно, широко, рассеянно или редко;
- распределение многообразия форм таксона в пределах ареала;
- положение по отношению к ареалам близкородственных таксонов;
- географическое положение ареала.
Путем сравнительной оценки многочисленных ареалов с этих точек зрения могут быть установлены различные типы ареалов (см. рис. 14.14).
- Протяженность ареалов
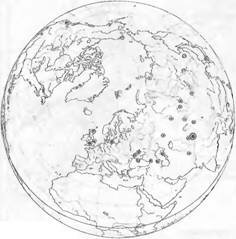
© Верхняя юра ® Средняя юра © Нижняя юра
Рис. 14.16. Распространение рода Ginkgo в Северном полушарии по ископаемым находкам от нижней юры до раннего плиоцен-неогенового периода и нашего времени (по H.Tralau).
В основе карт лежат современные очертания земной поверхности. Изменения с начала юрского периода (почти 200 млн лет назад) оказывали незначительное влияние на потенциальные пути миграции в Северном полушарии
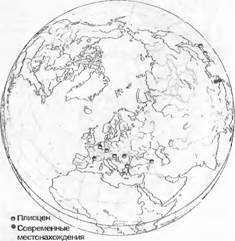
Между таксонами, произрастающими только в одном местонахождении в мире, и теми, которые в пределах своей приуроченности к определенному местообитанию произрастают по всему миру, имеются все возможные промежуточные формы.
Таксоны, ограниченно произрастающие в пределах единственного, как правило, небольшого ареала, называются эндемичными, причем это понятие используется довольно неопределенно. Наряду с действительными редкостями, в основном на видовом уровне локальными эндемиками, различают также региональные (например, произрастающие только в Альпах) или даже континентальные эндемики (например, произрастающие только в Австралии), причем последняя категория имеет смысл только на уровне семейства. Есть эндемики старые, реликтовые — палеозлдемики и молодые — неоэндемики. Широко распространенные космополиты — это преимущественно растения, произрастающие в условиях, связанных с деятельностью человека, но есть и природные космополиты, прежде всего среди споровых растений.Из известных реликтовых эндемиков, которые когда-то были распространены широко, можно назвать Ginkgo biloba (сейчас он еще растет только в западной части Китая; рис. 14.16), Sequoiadendron giganteum (Калифорния) и Wel- witschia mirabilis (юго-западная Африка). В противоположность им эндемичными ареалами обладают Betula oycoviensis (юго-восточная Польша), Papaver kerneri (юго-восточные Доломитовые Альпы); виды рода Erysimum sect. Cheinmthus в Эгейской области (см. рис. 10.32), как известно. возникшие в результате скрещивания, — это расположенные рядом ареалы близкородственных видов, все они относятся к относительно молодым эндемикам (новым видам). Доля эндемичных таксонов, очевидно, увеличивается с возрастом и степенью изоляции жизненного пространства.
В качестве примеров космополитных видов среди споровых растений можно привести печеночный мох Marchantia polymorpha или папоротник Pteridium aquilinum, болотные и водные растения, широко распространяющиеся водными птицами: водяную сосенку (Hippuris vulgaris), тростник (Phragmiles australis), а также многочисленные, распространяемые человеком сорняки, которые на нарушенных участках (рудеральних местах, часто у обочин дорог) можно найти по всему земному шару (например, вилы родов Plantago, Роа, Rumex, Senecio, Stellaria, Trifolium).
На уровне семейств по всему миру встречаются те, которые по своей природе имеют высокое видовое богатство (Orchidaceae, Poaceae, Asteraceae, Fabaceae).- Естественные разрывы ареалов
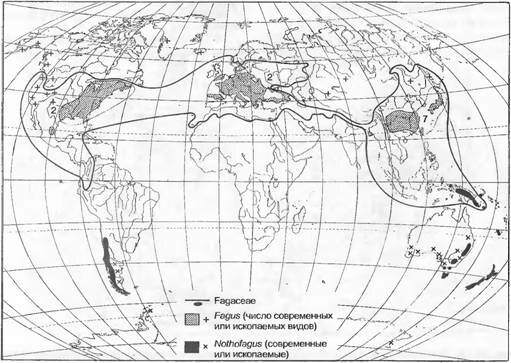
Рис. 14.17. Современное общее распространение семейства буковых (Fagaceae) с ареалами рода Fagus и родственного ему семейства Nothofayaceae с ареалами рода Nothofagus, а также ископаемые находки (+, х) обоих родов (по H.Meusel et al.; С.G.G.J.van Steenis)
Полностью непрерывное заселение встречается редко. По меньшей мере по направлению к окраинам ареалов распространение большинства таксонов рассредоточивается, ограничиваясь отдельными форпостами (или, наоборот, местонахождениями, оставшимися при отходе). Если пробелы в ареалах оказываются такими большими, что таксоны не могут их преодолеть обычными способами распространения, говорят об эксклавах и, соответственно, дизъюнкциях. Многие дизъюнктивные узоры повторяются довольно регулярно (генетические последствия искусственного фрагментирования местообитаний, см. 10.3.2.2).
У Pinas sylvestris на севере мы видим относительно сплошной основной ареал, в южных же горах имеются дизъюнктивные, более мелкие частичные ареалы, или эксклавы. Почти одинаковы по протяженности три дизъюнктивных частичных ареала рода Fagus (рис. 14.17). Тот факт, что дизъюнкции у Fagus, Hepatica (см. рис. 14.17) и других родов, свойственных широколиственным лесам, очень сходны, объясняется общими историческими и климатическими причинами. При наличии дизъюнкций можно думать или о редукции сплошной в прошлом области распространения или, в чрезвычайных случаях, о привнесении из очень далеко расположенной области. Для гибридогенных таксонов (например, у аллополиплоидов) или экологических рас нельзя исключить возможности неоднократного параллельного возникновения на разных местах (политопное происхождение).
- Плотность заселения ареалов

'T/I?. • Ononis (с числом видов) 5Ж а 10 видов gt; 30 видов
Рис. 14.18. Центр разнообразия и зоны снижения числа (обозначены цифрами) видов для рода Ononis (стальник, Fabaceae) (по H.Meusel, E.Jager)
Высокая плотность заселения указывает на то, что для данного вида область экологически оптимальна. При этом часто достигаются более сильная его экспансия на различные местообитания (максимальная экологическая амплитуда) и наибольшая генетическая диверсификация (многообразие форм, центр вариабельности). Аналогично и у родов или семейств, для которых регионы с особенно большим видовым богатством могут считаться центрами разнообразия. Это также области процветания и соответственно поддержания таких таксонов, но они не обязательно представляют собой центры происхождения. По всем этим критериям можно установить так называемое ядро, или центр ареала.
Очанка крошечная (Euphrasia minima) произрастает в Центральных Альпах от горного до нивального пояса (1 400—3 100 м); ее максимальная встречаемость, наиболее широкая экологическая амплитуда и центр варьирования приходятся на нижний альпийский пояс между 2000 и 2 300 м (49 фенотипов по сравнению с двумя на верхней и восьмью на нижней границах распространения). Пример центра разнообразия рода — род Ononis, представленный наибольшим количеством видов в юго-западном Средиземноморье (рис. 14.18), для семейства— Rubiaceae (Мареновые), с высоким родовым разнообразием в теплых и влажных тропиках и прогрессирующим его убыванием при переходе к более сухим или холодным областям.
Еще по теме Типы ареалов:
- ТИПЫ АРЕАЛОВ[66]
- ТИПЫ АРЕАЛОВ
- Ареал
- АРЕАЛ
- Метод типизации ареалов
- Представления континуалистов об ареале
- Географические связи между ареалами
- Методы изображения ареалов и проведения их границ
- Причины, обусловливающие границы и площадь ареалов
- Глава XII АРЕАЛЫ АККУМУЛЯЦИИ ПРОДУКТОВВЫВЕТРИВАНИЯ И ПОЧВООБРАЗОВАНИЯ
- 2. 1. 4. Другие факторы, влияющие на ареал вида
- Глава IV АРЕАЛ И БИОГЕОГРАФИЧЕСКОЕ ДЕЛЕНИЕ
- Представления континуалистов и структуралистов об ареал
- Типы размножения Животные
- Глава 3. ИСТОРИЯ АККЛИМАТИЗАЦИИ ОНДАТРЫ И ЕЕ СОВРЕМЕННЫЙ АРЕАЛ
- 2.2. ТИПЫ КЛЕТОЧНОЙ ОРГАНИЗАЦИИ
- ТИПЫ ПИТАНИЯ РАСТЕНИЙ
- ТИПЫ ЛЕСА НА ОСУШЕННЫХ ТОРФЯНЫХ ПОЧВАХСЕВЕРНОЙ БЕЛАРУСИ