Причины, обусловливающие границы и площадь ареалов
Границы ареалов обусловлены не только историческими и физическими причинами (например, взаимного размещения материков и морей), но и многими экологическими, поскольку связаны с условиями современного климата и почвенным покровом (см.
рис. 12.5, 14.1), а также с экофизиологическими свойствами видов растений. Значительная часть ареалов обусловлена связанными с температурой зонами растительности или придерживается секторов, обусловленных океаничностью климата (см. 12.5.2.2). Многократно проводились попытки наложить границы ареалов на определенные изолинии климатических показателей, чтобы рассматривать последние как климатические факторы формирования границ (рис. 14.25). Ввиду комплексной природы климатических и почвенных условий, а также сочетания их с явлениями конкуренции подобные попытки тем не менее остаются проблематичными.Имеющие ступенчатую форму восточные границы распространения Fagus sylvatica (бука) и Quercus robur (дуба черешчатого) явно обусловлены климатически (см. рис. 11.259). Дуб значительно устойчивее к экстремальным температурам и засухам, чем бук. У атлантического падуба (Ilex aquifotium), имеющего близкородственные виды не только в западной, но и в восточной Евразии и в Гималаях (см. рис. 14.25), южная и восточная границы совершенно явно обусловлены возрастанием летней сухости. Суровость зим (континентальность!) также связана, но не напрямую, с указанными температурными кривыми. Решающими здесь очевидно являются низкие зимние температуры (менее -15 °С), вероятность произрастания коррелирует с этой изолинией, ощутимые повреждения морозами влекут за собой тяжелые последствия и обусловливают ослабление конкурентоспособности. Ареал Hex заметно сузился после экстремальной зимы 1928/29 гг.
Даже когда ареал вида из чисто прагматических соображений называют замкнутым, вид в его пределах всегда ограничен своими специфическими местообитаниями, причем размеры и частота встречаемости этих местообитаний определяют внутреннюю плотность заселения ареала.
Значительное перекрытие на плоской проекции ареалов разных таксонов вовсе не означает, что эти виды или группы видов имеют одинаковые требования к условиям среды. Ареал всегда неизбежно покрывает весь спектр типов местообитаний ландшафта, различных по положению в рельефе, экспозиции, влажности и субстрату.У бука, дуба и сосны в Центральной Европе ареалы значительно перекрываются. Однако в пределах одной области они заселяют тем не менее совершенно разные местообитания, поэтому часто не растут в одних и тех же сообществах, но являются характерными видами буковых, дубовых и сосновых лесов. Если сравнить определенные в чистых культурах примерные оптимальные значения влажности и pH почвы, то между этими тремя видами обнаружится очень мало различий. В смешанной же культуре светолюбивый дуб отступает от теневыносливого бука в краевую зону благоприятных для него по условиям мест, а еще более светолюбивая сосна почти полностью подавляется. Сосна ни в коем случае не «предпочитает» в Центральной Европе более сухие места, или сырые болотные, или участки с бедными и кислыми почвами, но она может выдерживать такие экстремальные условия, потому что там отсутствует конкуренция со стороны более требовательных к условиям лиственных пород (см. 12.1).
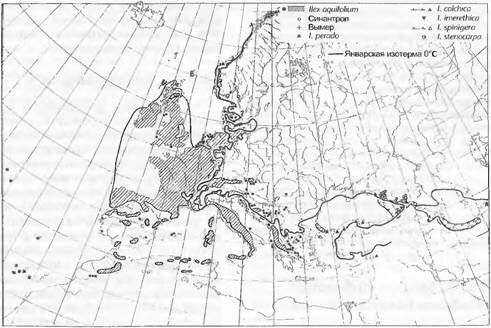
Рис. 14.25. Распространение падуба {Ilex aquifolium) и близкородственных видов рода в западной части Евразии.
Для сравнения приведены январская изотерма 0 °С по Н. Meusel et al. и изотерма по Климатическому атласу Европы (Юнеско, 1970)
Некоторые виды отличаются удивительно широкой амплитудой по отношению к климату и в пределах ареала находятся в самых различных климатических зонах, что часто наблюдается вместе с явственной экотопической дифференциацией популяций. Хороший пример из этой области имеется также для лесообразующих пород: ареал сосны простирается от Южной Испании до Лапландии, бука — от Сицилии до Северной Скандинавии (см.
рис. 11.259). Это стало возможным, с одной стороны, благодаря экологической компенсации (на юге они занимают северные склоны вместо южных или поселяются на больших высотах), с другой — образованию различающихся экотипов (см. рис. 10.27, В). Fagus sylvatica в центральной части своего ареала распространен от пояса холмистых равнин до верхнего горного, но на юге он ограничивает свое распространение более холодными горными лесами, а на северной границе растет на почвах низменных равнин. Еор- ные растения, например осока твердая (Carex firma) или растущая в поясе криволесья горная сосна (P'mus mugo), могут часто селиться в условиях холодно-влажного местного климата даже на равнинных поверхностях.Часто на основании локального распространения вида и его связи с местообитанием можно судить о его общем распространении и наоборот; то и другое, являясь как бы «общим знаменателем», лежит в основе экофизиологической шкалы потребностей и допустимых пределов существования вида или группы видов (часто описывается как норма реакции). А потому карты распространения можно было бы употребить при соответствующей критической оценке как показатель определенного фактора местообитания. В какой мере они отражают причинные связи, вопрос масштаба. Мелкий масштаб дает общее представление о требованиях к макроклимату. Более крупный масштаб с большей вероятностью отражает эдафические причины, обусловливающие распространение. Связь между ареалом и современными условиями окружающей среды особенно бросается в глаза на примере видов, жестко привязанных к определенному типу почвы. С давних пор известны растения засоленных, песчаных, известковых и щебнистых
грунтов/см. 6.2.2.4, 13.6.6).
?
Как пример, сошлемся здесь на солевыносливые (например, Salicornia europaea agg., Glaux maritima и Aster tripolium) и предпочитающие пески (например, Salsola kali и булавоносец, Corynephorus canescens) растения морских побережий и удаленных от моря участков. Только на серпентиновых скалах произрастают некоторые папоротники, например Asplenium adulterinum. В Альпах встречаются лишь на известняковых или соответственно на силикатных породах близкие, но не близкородственные (псевдовикари- руюшие, см. рис. 14.14, Е) пары видов Rhododendron hirsutum и R. ferrugineum, Gentiana clusii и G. acaulis s. str. (= G. kochiana). При этом решающую роль играет реакция почвы (см. 6.2.3). Среди полевых сорняков дикая редька, Raphanus raphanistrum, растет на более или менее кислых почвах, а полевая горчица, Sinapis arvensis, напротив, на основных или очень слабокислых. Постоянство по отношению к почвам вообще может меняться со сменой климатических условий; многие виды, которые в более сухих и теплых районах растут на различных почвах, в районах с холодным и влажным климатом становятся кальцефилами, т.е. здесь та же основная или нейтральная реакция почвы имеется только на известняках.
Еще по теме Причины, обусловливающие границы и площадь ареалов:
- Методы изображения ареалов и проведения их границ
- ФАКТОРЫ, ОБУСЛОВЛИВАЮЩИЕ БИОТЕРМИЧЕСКОЕ ОБЕЗВРЕЖИВАНИЕ НАВОЗА
- ПЛОЩАДЬ ПИТАНИЯ, ПРОРЕЖИВАНИЕ ВСХОДОВ
- Ветровальная площадь
- СОКРАЩЕНИЯ ПЛОЩАДИ И БИОМАССЫ ЛЕСОВ
- Площадь листовой поверхности посевов кукурузы
- Метод типизации ареалов
- РАЗВЕРТЫВАЮЩАЯСЯ ПЕРМАКУЛЬТУРА ДЛЯ БОЛЬШИХ ПЛОЩАДЕЙ
- Ареал
- АРЕАЛ
- Границы между сообществами и ценотическими системами
- Представления континуалистов об ареале
- Метод значимости биогеографических границ
- Географические связи между ареалами
- Типы ареалов
- ТИПЫ АРЕАЛОВ[66]
- б. КОЛЕБАНИЯ ГРАНИЦ И ВЕРОЯТНОСТНЫЙ ПОДХОД К БИОТОПУ
- Проекты за границей
- НЕВИДИМЫЕ ГРАНИЦЫ