Представления континуалистов об ареале
«Возникновение вида происходит в одном месте, откуда он начинает завоёвывать пространство... Это положение является ключевым для понимания истории формирования ареала» (Петров, 2001: 47).
Согласно континуалистам, ареал вида определяют свойства самого вида. Вид возникает в каком-либо месте, откуда со временем распространяется, т. е. в момент возникновения имеет очень маленький ареал. Сторонники христианской традиции сначала считали, что все виды животных и растений (по крайней мере наземных) распространились из одной точки — г. Арарат, поэтому эту концепцию по приоритету следует называть араратской. По мере накопления фактов, одна точка расселения для всей наземной биоты была заменена на несколько (но для каждого конкретного вида точка возникновения-расселения по-прежнему предполагалась одна). Такое представление о динамике ареала нашло своё выражение в формуле ageamp;area: «чем старше таксон, тем шире его ареал» (Willis, 1922). Исходя из этих представлений, если ареал какого-то вида мал, то он или ещё не успел расселиться (неоэндем), или ещё не успел вымереть (палеоэндем). Я.И. Старобогатов (1970) даже пытался использовать дисперсионный и корреляционный анализ для оценки роли исторических факторов в размерах площади ареалов видов, попытка эта мало обоснована и наивна.
Наилучшим подтверждением этой теории были бы палеонтологические данные, но их по отдельным видам практически нет. Поэтому, несмотря на большую популярность араратской теории, весьма трудно, если вообще возможно, привести пример нового вида, только что возникшего и увеличивающего площадь своего ареала. Многочисленные примеры увеличения ареала относятся к (1) видам, которым человек помог преодолеть существующие барьеры или к (2) ценофобам, расселяющимся по сообществам, модифицированным человеком в той или иной степени. Часто вид относится к обоим группам одновременно.
Данные же по более крупным таксонам весьма противоречивы. Наряду с большим количеством данных, подтверждающих араратскую теорию, имеются и данные, противоречащие ей. Так, А.Н. Миронов, исследуя распространение морских ежей в Мировом океане, обнаружил, что «в семействах Pourtalesiidae и Urechinidae чем позднее возник род, тем более широким географическим и вертикальным распространением он характеризуется» (Миронов, 1982: 489).
Но всё же данные по надвидовым таксонам не вполне подходят для изучения динамики ареала отдельных видов, поэтому были предприняты усилия по сбору информации, подтверждающей временнэю динамику видовых ареалов. Одним из типов данных, вроде бы подтверждающих араратскую теорию на видовых ареалах, являются многочисленные случаи, показывающие уменьшение плотности вида по направлению к краям его ареала. Их интерпретируют как свидетельство существования оптимума ареала вида, где он заселяет наибольший спектр местообитаний, где постоянно или периодически наблюдают максимальную для вида плотность поселений. По направлению от оптимума к периферии ареала его обилие падает, что интерпретируют как непрерывно происходящий процесс расселения вида из оптимума его ареала до достижения видом условий, выходящих за рамки его экологической амплитуды, где он перестаёт размножаться и образует стерильные или зависимые поселения.
Такие данные обычно получают на основе континуалистской парадигмы: путём подсчёта какого-либо показателя (например, частоты встречаемости) в каких-либо априорно выделенных площадях (например, квадратах фиксированного размера) без учёта подходящих для вида сообществ и тем более без учёта границ ЦС. Такой подход в значительной степени предопределяет и результат, иными словами, изменение обилия вида от центра его ареала к периферии в значительной степени может быть артефактом применённых методик сбора и обработки материала, а не отражением реальной ситуации. И в первую очередь это касается ценофильных видов.
Любой ценофильный вид — компонент ассоциации и вне неё не встречается.
Невозможно обнаружить дуб посреди озера, а душистый колосок в дубраве, хотя при продолжительности наблюдений, равной продолжительности протекания сукцессии мы зарегистрируем дубы на месте ранее существовавшего озера, а луг из душистого колоска возникнет на месте дубравы после вырубки. Обычно в силу чисто случайных по отношению к ЦС причин (неоднородность рельефа, гидрологии и т. п.) одни сообщества занимают булыпую долю площадей в одной части ареала ЦС, другие — в противоположной. При должном усердии, как правило, удаётся подобрать фактор, дающий нужный градиент в нужном направлении.Кроме того, на границе ареала вида, в анализируемые площади неизбежно попадут участки за пределами ареала вида. Усреднение таких данных даст снижение всех показателей по краям ареала.
Таким образом, изначальный выбор подхода, как это часто бывает, предопределяет ответ, не выявляя действительную картину распределения вида по ареалу.
Проверка представлений об уменьшении плотности поселений вида к границам ареала на некоторых массовых видах полихет Северного Ледовитого океана показала, что ни в одном из случаев справедливость этого мнения достоверно не подтверждается. Более того, в некоторых случаях это мнение достоверно несправедливо. Так, один из наиболее обычных и массовых видов полихет Северного Ледовитого океана, широко распространённый также и в Пацифике (от Берингова пролива до Фолклендов и Японии) — Nothria hyperborea — является массовым видом в Норвежском море вплоть до самых границ своего ареала (рис. 5.1), где резко исчезает на очень короткой дистанции (судя по имеющейся довольно редкой сетке станций — не более первых десятков километров).
«На крайнем северном пределе своего распространения полёвка-экономка нередко достигает плотности поселения, которая характерна для лесостепных популяций (экологический оптимум вида) в период пика численности» (Шварц, 1980: 219).
Аналогичные примеры известны и из других таксонов. Так, ель Piceci cibies у самой южной границы своего ареала в Московской области у устья Лопасни образует обширные ельники (Алехин, 1947), в чём каждый легко может убедиться, о каком-либо угнетении отдельных деревьев, как должно бы быть по араратской теории, не может быть и речи.
Далее континуалисты полагают, что в пределе вид увеличивает свой ареал до тех пор, пока не встретит условия, при которых он существовать не может. Сторонники такой весьма популярной точки зрения даже не допускают ситуации, в которой вид не встречается в данном регионе, хотя и может туда попасть, и имел достаточно для этого времени, а абиотические условия являются для него подходящими. Так А.Г. Воронов (1963) и А.Ф. Алимов с соавторами (2004), упрощая классификацию границ В.В. Станчинского (1922) с 4 типов до 2, отбрасывают ривалитатные (зависящие от биотических причин) границы, которые сам В.В. Станчинский считал столь же значимыми, как и границы, обусловленные действием абиотических факторов (с точки зрения структурализма не ривалитатных границ вовсе нет), с чем были согласны даже географы (например, Л.С. Берг).
Значения абиотических условий, ограничивающих распространение вида, естественно, различны для разных видов, поэтому границы потенциальных ареалов отдельных видов могут совпадать только случайно. Если бы градиент абиотических условий на поверхности Земли был бы постоянен, то границы ареалов видов распределялись бы случайно. В действительности наблюдается чередование областей с резкими и слабыми градиентами. Области резких градиентов являются аттракторами границ (см. предыдущую главу), поэтому на поверхности Земли чередуются области сгущения и разрежения границ ареалов. Это чередование можно обнаружить статистическими методами. Континуалист- ская биогеография считает, что причиной появления областей сгущения границ ареалов является наличие областей с резкими градиентами абиотических факторов и свою основную задачу видит в выявлении таких областей.
Я.И. Старобогатов (1970: 91) писал: «Фактические ареалы будут тем меньше потенциальных, чем большее значение в распространении данного вида играют исторические факторы», под которыми он понимает процесс расселения вида из места его возникновения. «Районирование, построенное на легко расселяющихся организмах, будет менее дробным, чем построенное на организмах, обладающими малыми способностями к расселению» (Старобогатов, 1982: 16).
Разбирая эту умозрительную гипотезу, прежде всего надо отметить, что предположение о прямой связи размера ареала с лёгкостью расселения отнюдь не всегда подтверждается фактическими данными.Представление о влиянии малых способностей к расселению на размер ареала основано на смешении двух совершенно разных явлений: способности к расселению самой по себе и способности к преодолению преград. В качестве примера рассмотрим распространение полихеты Anobothrus sp. nov. (рис. 5.6). Хотя биология размножения этого вида неизвестна, он принадлежит к семейству, все изученные виды которого не имеют пелагических личинок, поэтому с высокой вероятностью можно считать, что пелагической личинки нет и у него. Этот вид — стенобатный абиссальный, известен с глубин 3460-8050 м, поэтому проникновение его в Северную Атлантику из Северной Пацифики через Север-
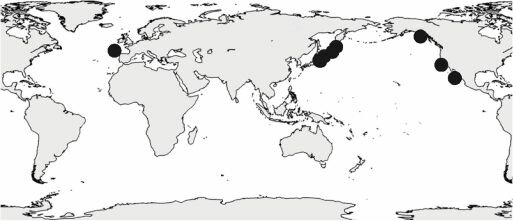
Рис. 5.6. Ареал Anobothrus sp. nov.
современная граница ареала дуба
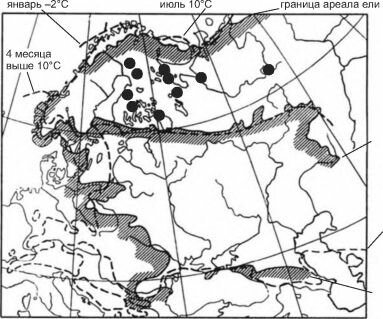
январь +2°С
граница ареала бука
Рис. 5.7. Северные границы ареалов ели Picea abies и дуба Quercus robur и восточная — бука Fagus sylvatica в сопоставлении с клималиниями (прерывистые линии). По Вальтер, 1982).
Кружками показаны нахождения дуба 2500 лет назад. По С.А. Турубановой (2000).
ный Ледовитый океан исключено. Этот ареал и многочисленные аналогичные примеры однозначно свидетельствуют об отсутствии связи размера ареала вида с его способностью к расселению в пределах ЦС, в которые он входит. Вероятно, это можно объяснить тем, что за время существования ЦС любой вид со сколь угодно малыми способностями к расселению способен расселиться по всей её площади. Другое дело, если вид в своём распространении встречает преграды.
В этом случае способность к их преодолению действительно зависит от способности вида к расселению. Многочисленные примеры этой связи можно найти в ареалах пресноводных беспозвоночных и биогеографии островов.Для подтверждения того, что границы ареалов видов определяют абиотические факторы, континуалисты приводят карты ареалов, на которые нанесены изолинии абиотических факторов (клималинии), более или менее совпадающие с границей ареалов. Это совпадение трактуют как причинно-следственные отношения. Примером такой карты является карта из Г. Вальтера (1982, воспроизведена также: Абдурахманов и др., 2003: 151) (рис. 5.7). На мой же взгляд, самое поразительное на этой карте то, что даже столь квалифицированный учёный, как Г. Вальтер, не смог подобрать и трёх видов, распространение которых подтверждало бы справедливость его мнения. В самом деле, если совпадение границ ареалов дуба и ели с клималиниями можно считать хорошим (по крайней мере на изображённых на карте отрезках границ), то совпадение границы бука — весьма сомнительно, реально с клималинией совпадают только незначительные участки в Крыму и Румынии, а расхождение на Украине достигает нескольких сотен километров.
Не удивительно, что легко можно подобрать и противоположные примеры, когда вид прекрасно себя чувствует далеко за пределами своего ареала. С позиции континуализма совершенно необъяснимо, почему он там не встречался ранее. На рис. 5.8 приведён естественный ареал конского каштана. Вполне вероятно, что можно подобрать какой-либо показатель абиоты, изолинии которого совпадут с границами его естественного ареала. Однако, достаточно выглянуть в окно Биофака МГУ, или пройтись по улицам украинских или южнорусских городов, чтобы убедиться, что этот вид прекрасно себя чувствует и во-
«О Я.И. Френкеле рассказывают, что якобы в ФТИ в 30-е годы его изловил в коридоре некий экспериментатор и показал полученную на опыте кривую. Подумав минуту, Я.И. дал объяснение хода этой кривой. Однако выяснилось, что кривая случайно была перевернута вверх ногами. Кривую водворили на место и, немного поразмыслив, Я.И. объяснил и это поведение кривой» (Березинский, 1993: 16).
зобновляется и в совершенно других условиях (я сам видел у Биофака подрост возраста не менее 7 лет, но его регулярно скашивают и постоянно вытаптывают). У меня на участке на северо-востоке Костромской области выращенные из семян конские каштаны перенесли мороз в -43°С. Многочисленные аналогичные примеры даёт практика ботанических садов мира. Невозможно с позиций кон- тинуализма объяснить и многие из примеров, приведённых в разделе «Внедрение видов».
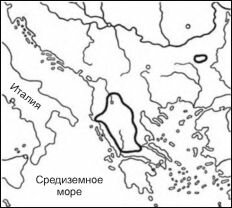
Рис. 5.8. Естественный ареал конского каштана Aesculus hippocastanum. По Шаферу (1956).
Число подобных примеров легко можно увеличить. Приведу только один: распространение уже упоминавшейся полихеты Nothria hyperborea (рис. 5.9). Этот вид в массе встречается на шельфе и склоне Тихого и Северного Ледовитого океанов и северо-западной Атлантики (местами он даже является руководящим видом сообществ), но далее на юг в Атлантике не идёт. Легко можно подобрать изолинию какого-либо абиотического фактора, которая объясняла бы такое положение южной границы его ареала, например, температуры. Но только в Атлантике. Распространение же вида в Пацифике никак не будет соотносится с аналогичной изолинией, что свидетельствует о том, что и в Атлантике это совпадение случайно.
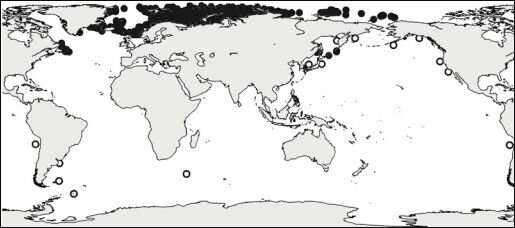
Рис. 5.9. Ареал полихеты Nothria hyperborea.
Светлые кружки по Н.В. Кучеруку (1985, в оригинале на рисунке вид по вине редакторов ошибочно назван Nothria solenotecton), тёмные кружки — по нашим данным.
Представления структуралистов об ареале
«Рассматривая современное распространение серии близкородственных клёнов ряда Picta, в частности встречающихся и в СССР: Acer laetum (Кавказ, горы северного Ирана и северо-востока Малой Азии), A. turkestanicum (горы Средней Азии), A. mono (Приамурье, Северо-Восточный Китай, Корея) и A. pic- tum (Сахалин, Япония), мы едва ли можем себе представить эти виды иначе, как занимающими весь современный ареал каждого из них с самого начала географической дифференциации когда- то широко распространённого исходного типа «laetum-pictum». Более того, принимая во внимание дизъюнктивный характер современного распространения этой группы видов, скорее можно предполагать, что ареал каждого из них во время их становления мог быть даже несколько шире современного» (Толмачев, 1974: 47).
Согласно структуралистам, ареал вида определяют свойства ЦС, в которые он входит. В момент возникновения вид уже может иметь ареал, существенно отличный от точки. Дальнейшую динамику ареала определяют разные факторы в случае ценофильных и ценофобных видов.
Ценофилъные виды. Потенциальные ареалы ценофильных видов с одинаковыми ареалами могут совпадать, а могут не только не совпадать, но даже весьма сильно отличаться от границ их фактических ареалов. Ареал вида в момент возникновения совпадает с ареалом формирующейся ЦС = биогеографического региона.
Расширение ареала ценофильного вида происходит следующим образом:
«В природный условиях лишь немногие виды можно рассматривать как изолированные популяции, потому что отдельные виды в большинстве случаев представляют собой взаимодействующие части многовидовых систем» (Джиллер, 19X8: 9).
«Экология может в принципе ответить иа вопрос, что обусловило то или иное географическое распространение вида. Но она обычно не может ответить на вопрос, почему вида нет там, где для этого действительно имеются все необходимые условия» (Беклемишев, 1982: 10).
- В норме расселение ценофильных видов происходит не самостоятельно, а постольку, поскольку расселяется вся ЦС. К совершенно аналогичному выводу пришел на основании изучения ареалов А.И. Толмачев, полагавший, что происходит «сопряжённое расселение целых серий таких ценотически взаимосвязанных видов, образующих на новом субстрате «привычные» биоценотические сочетания» (Толмачев, 1974:
211).
- При включении ценофильного вида в другую ЦС он увеличивает свой ареал на ареал этой ЦС.
Т. о., ареал ценофильного вида равен сумме ареалов ЦС, в которые он входит. Говоря об ограничении распространения ценофильных видов границами ЦС, в которые они входят, я не имею в виду невозможность расширения ареала ценофильного вида. Расселение вида возможно, более того, часто происходит, но вероятность того, что он закрепится за пределами ареала, низка, об этом свидетельствует сам факт совпадения границ ареалов ценофильных видов друг с другом и с границей ЦС, в которые они входят.
Фактически парадигма структурализма даёт биогеографу очень мощный способ экстра- и интерполяции при установлении ареала вида. Поскольку распространение любого ценофильного вида определяется распространением ЦС, частью которой он является, то индикатором при экстраполяции может служить любой другой ценофильный вид. Ещё лучшим индикатором является вся совокупность ценофильных видов, поскольку её распространение известно лучше, чем любого из них в отдельности. На практике можно использовать не всю совокупность, а достаточно большую выборку.
Ценофобные виды. Их ареал соответствует представлениям континуалистов поскольку его определяет только фундаментальная ниша вида. Однако это соответствие касается только границ ареала, а не распределения вида внутри ареала. Оно может соответствовать, а может и резко отличаться от представлений континуалистов. Примером такого принципиально иного распределения является распределение некоторых пограничных видов (Семёнов, 1982), которые обитают только вблизи границ ЦС.
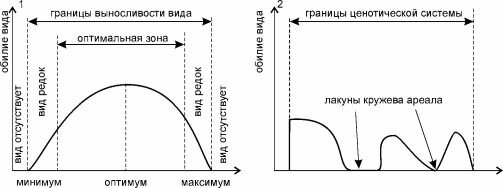
пространство пространство
Рис. 5.10. Распределение обилия вида в пределах его ареала.
1 — с точки зрения континуалистов; 2 — ценофильного вида с точки зрения структуралистов (для простоты рассмотрен вид, эндемичный для одной ЦС).
Вид является ценофобным или ценофильным только в пределах конкретной ЦС. Известны случаи, когда вид является ценофильным в одной ЦС и ценофобным — в другой. Ареалы таких видов совмещают свойства ценофильных и ценофобных (в разных своих частях).
При вселении ценофильного вида в ареал другой ЦС как ценофобного, вид постепенно увеличивает свой ареал до достижения пределов его потенциального ареала, т. е. границы ареала определяет фундаментальная ниша вида или проще говоря, его биология.
Схематически точки зрения континуалистов и структуралистов изображены на рис. 5.10. Обратите внимание на то, что, в отличие от предыдущей главы (рис. 4.1 и 4.2), по оси абсцисс в обоих случаях отложено пространство.
Безусловно, в каждом конкретном случае можно подобрать «объясняющую» post factum причину — экологический фактор, значения которого ограничивают распространение вида, но прогностическая ценность такого объяснения невелика: такой фактор и его значения обычно приходится подбирать не только для каждого вида, но и для разных отрезков границы ареала одного и того же вида. С другой стороны, факт весьма точного совпадения в пространстве границ ареалов видов с самой разной биологией и не связанных явными биотическими взаимоотношениями, свидетельствует о наличии структуры, организующей распределение видов по поверхности Земли — ЦС, которая в первую очередь и контролирует распространение видов в географическом пространстве. Выявив границы ЦС, мы с высокой точностью сможем провести границы ареалов ценофильных видов, входящих в эти ЦС.
Некоторые примеры распределения видов в пространстве, соответствующие представлениям структуралистов, приведены ранее, дополнительные примеры любой может получить, глядя в окно поезда дальнего следования.
Еще по теме Представления континуалистов об ареале:
- Представления континуалистов и структуралистов об ареал
- АРЕАЛ
- Ареал
- Метод типизации ареалов
- Типы ареалов
- ТИПЫ АРЕАЛОВ[66]
- Современные представления о строении физической материи
- Географические связи между ареалами
- РАЗВИТИЕ ПРЕДСТАВЛЕНИЙ ОБ ЭКОСИСТЕМАХ
- Методы изображения ареалов и проведения их границ