Флюктуаиионная и сукиессионная динамика популяций
Обычно по годам численность любой популяции неодинакова. Такие автономные погодичные колебания численности С.С. Четвериков (1926) назвал «волнами жизни*. Теперь чаще используется термин популяционные волны.
Как было показано в разделе 9.1, у популяций различают флюктуационную динамику, которая имеет обратимый характер, и сукцессионную динамику направленного необратимого характера, когда многие популяционные параметры изменяются по ходу сукцессии фитоценоза.
Флкжтуационные изменения могут быть кратковременными или долговременными, носить резкий катастрофический характер или быть плавными, сопровождаться глубокими или лишь незначительными изменениями многих параметров популяции, вызываться внешними по отношению к популяциям причинами или вытекать из сущности внутрипопуляционных процессов. Продолжительность флкжтуационных динамических смен может составлять от 3—5 лет до 300 лет и более. В значительной степени это зависит от жизненной формы растений и реализуемой ими стратегии. У однолетников с г-стратегией флюктуации краткосрочные, а у Я-стратегов охватывают большие промежутки времени. Б.П. Колесников (1968) показал, как на протяжении жизни одного поколения лесообразующего древостоя происходят существенные изменения в нижних ярусах леса. Многие варианты флюктуаций описаны в работе JI.A. Жуковой (1995).
При флюктуационной динамике популяции в ней периодически изменяются многие параметры: возрастной, онтогенетический и виталитетный состав, численность особей и плотность популяции, иногда сужаются или расширяются размеры популяционного поля.
Как теоретическое обобщение явления флюктуаций выступает концепция волнового развития популяций А. А. Уранова (1975). А. А. Уранов различал малые волны, когда в популяции с определенной периодичностью поступают новые диаспоры, и большие волны, связанные с более полными перестройками популяций.
К сожалению, за последние 30-35 лет существенного вклада в развитие этой концепции не было внесено, и она ждет дальнейших исследований. Это связано с тем, что многие специалисты по популяционной экологии работают маршрутными методами, а изучение волнового характера динамики популяций требует многолетних стационарных наблюдений.Сукцессионная динамика популяций всегда имеет направлен ный характер и представляет собой необратимое изменение свойств особей и особенностей популяций, сопряженное с ходом сукцессии фитоценоза, в котором находятся данные популяции. В зависимости от адаптируемости популяций к сукцессионным изменениям растительного покрова в одних случаях сукцессионная динамика ведет к увеличению численности особей в популяции, повышению популяционной плотности, возрастанию виталитета популяции, а в других случаях в ходе сукцессии растительного покрова все популяционные изменения имеют противоположный характер. Тогда популяции уменьшаются в размерах, фрагментируются и в конечном итоге могут полностью выпадать из растительного сообщества.
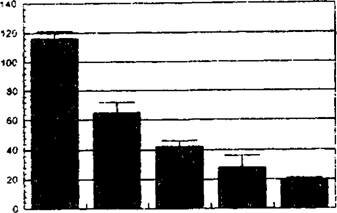
Распространенным вариантом сукцессионной динамики растительности является антропогенная динамика, в ходе которой популяции претерпевают наиболее глубокие и быстрые изменения. JI.H. Бондарева (2005) изучила ряд популяционных параметров ежи сборной Dactylis glomerata и щучки дернистой Deschampsia cespitosa в ряду пастбищной дигрессии, который был подразделен на пять ступеней. В зависимости от биолого-эколо- гических свойств этих двух видов растений динамические изменения популяций в этом случае были разнонаправленными (рис. 9.4). У ежи сборной Dactylis glomera ta от контрольного участка без выпаса до пятой ступени пастбищной дигрессии запас надземной фигомассы снижался почти в пять раз, в онтогенетическом спектре за счет повышения доли субсенильных и сенильных растений увеличилась правосторонность, а в виталитетном спектре господство в популяции особей виталитетных классов а и 6 сменилось абсолютным преобладанием угнетенных растений класса с.
В противоположность этому у устойчивого к выпасу растения щучки дернистой Deschampsia cespitosa (рис. 9.5) по градиенту пастбищной дигрессии запас фитомассы вырос почти в 7 раз,БКЯ ГД2 ГЩЗ ПД4 П?)5
Градиент пастбищной дигрессии
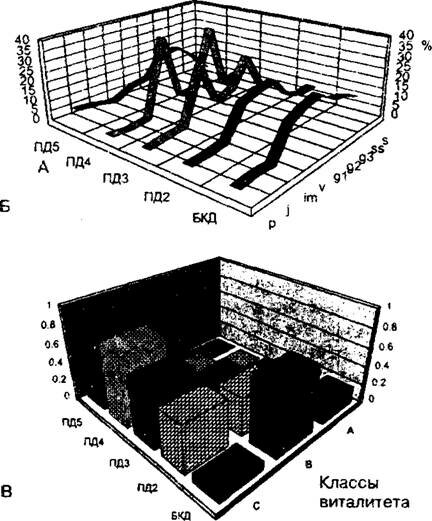
Рис. 9.4. Динамика популяционных параметров ежи сборной в ряду пяти фитоценозов пастбищной дигрессии:
А - снижение запаса фитомассы в популяции в граммах воздушно сухого веса на 1 м2; Б - изменение онтогенетического состава популяции; В - изменение виталитетного состава популяции (по Л.Н. Бондаревой, 2005 »
140
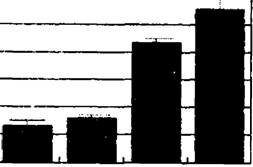
Рис. 9.5. Динамика популяционных параметров щучки дернистой в ряду пяти фитоценозов пастбищной дигрессии:
А - снижение запаса фитомассы в популяции в граммах воздушно сухого веса на 1 м2;

Б - изменение онтогенетического состава популяции;
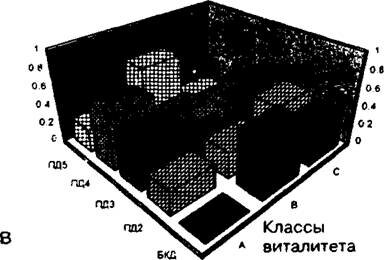
В - изменение виталитетного состава популяции(по Л.Н. Бондаревой, 2005)
онтогенетический спектр приобрел характер нормального, полночленного с преобладанием виргинильных и молодых генеративных особей, а в виталитетном спектре увеличилась доля растений виталитетного класса а.
Еще по теме Флюктуаиионная и сукиессионная динамика популяций:
- ЗАКОНОМЕРНОСТИ ДИНАМИКИ ЧИСЛЕННОСТИ ПОПУЛЯЦИЙ. МАТЕМАТИЧЕСКОЕ МОДЕЛИРОВАНИЕ ПОПУЛЯЦИЙ
- Математическое моделирование динамики популяций
- Компьютерное молелирование динамики популяций
- ДИНАМИКА ЧИСЛЕННОСТИ ПОПУЛЯЦИЙ
- 3. Динамика численности популяций
- Глава 9 Динамика популяций
- Динамика численности популяции во времени.
- § 1. РОЛЬ ДИНАМИКИ ВОЗРАСТНОЙ СТРУКТУРЫ ПОПУЛЯЦИИ В ПРЕОБРАЗОВАНИИ ЕЕ ГЕНЕТИЧЕСКОГО СОСТАВА
- ПРЕОБРАЗОВАНИЕ ПОПУЛЯЦИЙ. ГОМЕОСТАТИЧЕСКОЕ ИЗМЕНЕНИЕ ГЕНЕТИЧЕСКОЙ СТРУКТУРЫ ПОПУЛЯЦИЙ И микроэволюция
- Границы между популяциями, иерархия популяций
- Фазовый портрет динамики численности
- Динамики
- 7. Динамика численности
- Типы динамики численности
- Пятая динамика. Жизненные формы
- Четвертая динамика. Человечество
- Одиннадцатая динамика. Технология игры
- Динамика микробных комплексов
- Восьмая динамика. Бесконечность, Творец
- Глава 6. СЕЗОННАЯ ДИНАМИКА ПОЧВЕННЫХ ПРОЦЕССОВ