Термиты как разрушители растительных остатков
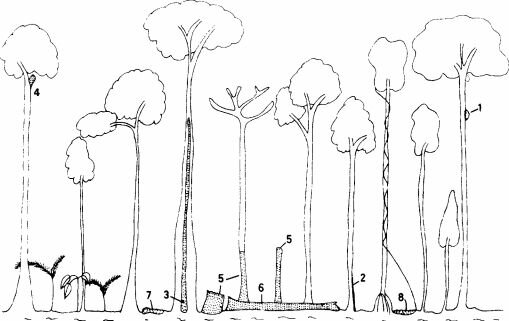
Рис.
2.1. Места, где термиты потребляют древесину в лесу Пасох-Форест, Западная Малайзия. /-гнезда вокруг стволов (например, Mic- rocerotermes); 2-поверхности вблизи оснований деревьев; 3-центральная часть ствола;4- отмершие ветки живых деревьев; 5-пни или неповалившиеся высохшие деревья; 6-повалившиеся стволы; 7-опавшие ветки и листья; S-гумус в почве. (Abe, 1979.)
Вполне возможно, что первоначально термиты были полифагами и питались разнообразным органическим материалом. Наиболее примитивный из современных видов Mastotermes darwiniensis, обитающий в Австралии,- полифаг; он поедает помет животных, растительные остатки и древесину. К полифагам относится и африканский термит Psammotermes hybostoma, который не брезгует почвой из подземных ходов и всевозможными отбросами. В выростах задней кишки у всех термитов обитают микроорганизмы, помогающие им переваривать пищу. У представителей наиболее примитивных семейств (Calotermitidae, Hodotermitidae и Rhinotermitidae) облигатные симбионты - жгутиковые простейшие - осуществляют внутриклеточное переваривание пищи; симбионты
Тermitidae - бактерии - переваривают целлюлозу. Задняя кишка термитов выстлана кутикулой, которая периодически сбрасывается, но благодаря тому, что особи обмениваются между собой пищей, реинокуляция обеспечена.
Это одно из важных преимуществ общественного образа жизни термитов.
И наши дни термиты играют важную роль в разложении мертвых деревьев в тропиках (рис. 2.1). Они очень разборчивы в пище. Например, в Австралии Nasutitermes exitiosus не нападает на один из видов Pirns, потому что содержащиеся в нем пинены служат феромонами тревоги для этого термита и их присутствие вызывает у него беспокойство.
Это может показаться невероятным, но многие другие виды поедают эту сосну, а N. exitiosus нападает на дерево с еще более сильным запахом-Eucalyptus marginata, которого большинство других термитов избегает.В
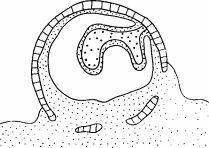
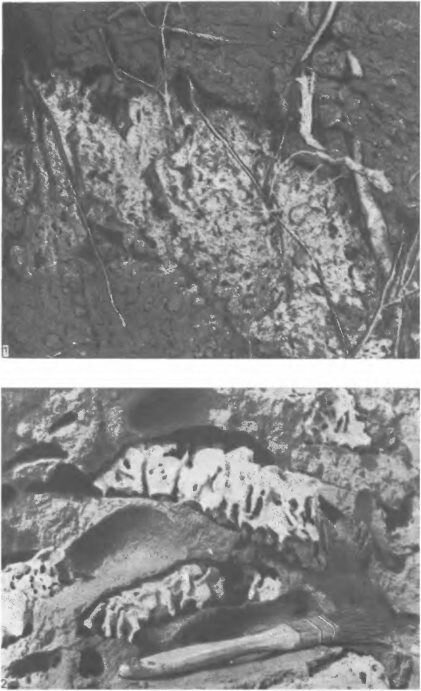
тельные количества почвы. В. Галереи расширяются, превращаясь в камеры, и бревно начинает погружаться в почву. Г. Бревно разрушается, термиты начинают покидать его, и оно разваливается. (Abe, 1980.)
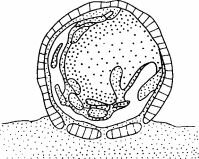
Рис. 2.2. Стадии разложения бревна. А. Термиты проникают через ходы, проделанные жуками, собирают почву и прилепляют ее к нижней поверхности бревна. Б. Термиты проделывают в бревне галереи и приносят дополни
Мягкие ткани термиты, по-видимому, предпочитают твердым (Wood, 1978); важное значение имеет также стадия разложения (рис. 2.2). Выделения грибов могут либо привлекать, либо отпугивать термитов в зависимости от их вида (Sands, 1969); при этом грибы иногда способны разрушать такие отпугивающие насекомых вещества, как скипидар. Привлекательность древесного метериала для термитов зависит также от его величины; например, в сухих лесах Южной Австралии N. exitiosus пожирает упавшие стволы и пни эвкалипта, а два других вида-только тонкие ветки. Большинство видов, питающихся древесиной, не трогают живые деревья, а многие явно предпочитают либо сухую древесину (Calotermitidae и др.), либо влажную (например, Termopsinae, сем. Hodotermitidae).
Древесина-это в основном смесь целлюлозы и лигнина, находящихся на разных стадиях разложения.
Целлюлоза - полимер глюкозы, а лигнин - полимер, состоящий из фенилпропановых единиц, образующихся из коричных спиртов. Соотношение лигнин/целлюлоза варьирует у разных видов в пределах от.
0,4 до 0,8. Calotermes flavicollis, принадлежащий к примитивному семейству Calotermitidae, переваривает 70-90% всей целлюлозы, тогда как Nasutitermes ephratae (сем. Termitidae) может использовать ее более чем на 90%.Лигнин переваривается гораздо хуже: не более одной трети у Calotermes flavicollis и около половины у Nasutitermes ephratae (La Fage, Nutting, 1978; Lee, Wood, 1971; Wood, 1978).
Представители подсемейства Termitidae (сем. Macrotermitinae) устраивают „сады“, в которых выращивают базидиальный гриб Termitomyces. По-видимому, нет сомнений в том, что эти сады устраиваются на частично переваренных экскрементах и что ни одно одиночное насекомое не могло бы создавать их. Показано (Josens, 1971а), что пища (древесные опилки с добавленной в качестве метки сажей) проходит через кишечник и откладывается термитами на поверхность сада; за 5-8 недель меченый субстрат опускается до основания сада, где поедается рабочими особями. Образующая сад масса содержит очень мало глиноподобного материала, находящегося в кишечнике рабочих особей, и она не похожа на их обычные экскременты. Симбиоз термитов с базидиомицетом, который растет в этой массе и разрушает здесь лигнин и целлюлозу, дает возможность полнее использовать волокнистые растительные остатки, собранные рабочими. Гриб
Фото 1. Грибной сад, выращиваемый в земляном гнезде муравья-листореза Асготугтех octospinosus. «Сад» представляет собой губчатую массу, на которую сверху добавляется новый субстрат, а муравьи объедают снизу нарастающие плодовые тела гриба. [D. I. Strad- ling.] (Здесь и далее в квадратных скобках ука- концентрирует питательные вещества в богатых белком клетках конидиеносцев, хитиновые стенки которых термиты легко переваривают-так же легко, как в случае надобности они переваривают кутикулу своих сотоварищей по гнезду, если их окажется слишком много. Хотя биохимическая сторона симбиоза в деталях еще не вполне ясна, нет сомнений в том, что гриб служит для термитов полезной добавкой к их пище (La Fage, Nutting, 1978; Rohrmann, Rossman, 1980) (см.
фото 1 и 2).Mac votennes michaelseni в Кении собирает вместо мертвой древесины траву, особенно в середине сухого сезона, в период роста половых особей. Термиты М. bellicosus в саванновом редколесье в Нигерии (Моква, 9° с. ш.) собирают ежегодно 177 кг/га древесины и 64 кг/га опавших листьев, что составляет примерно 6,4% общего количества лесного опада, образующегося за год. Термиты в целом собирают 24% (Collins, 1981a,b), но у Macrotermitinae уровень потребления пищи на единицу собственного веса вообще выше, чем у термитов из других групп; это, несомненно, обусловлено тем, что их „одомашненный“ гриб в отличие от „диких44 микроорганизмов активен на протяжении всего сухого сезона. Некоторые Hodotermitidae срезают сухую траву по ночам, выходя на повфхность, или днем, прячась под почвой. Это делают также представители двух подсемейств Тermitidae - Тermitinae и Nasutitermitinae. Hodotermes mossambicus в Южной Африке
заны лица, любезно предоставившие фотографии для публикации в настоящей книге.)
Фото 2. Грибной сад термита Macrotermes subhyalinus. «Сад» занимает одну из камер гнезда, где он располагается на маленьких конических подпорках (см. также фото 16). [J.P.E.C. Darlington.]
предпочитает траву, а не кустарники, и притом не свежую зеленую траву, а сухую.
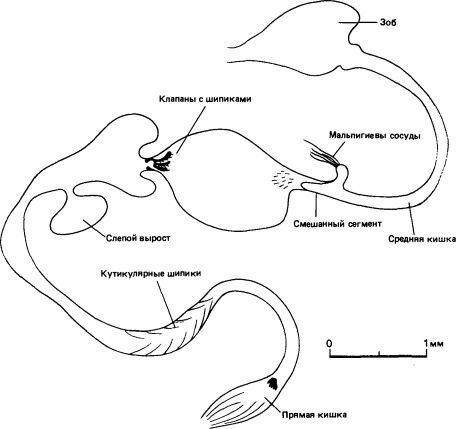
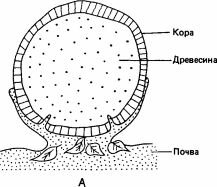
Рис. 2.3. Кишечный тракт термита Procubitermes. Показаны огромные размеры и сложность строения задней кишки, т. е. участка, расположенного позади мальпигиевых сосудов. (Bignell et al., 1980.)
Trhwrvitermes geminatus предпочитает злаки с нежными, а не грубыми листьями и отказывается от Cymbopogon, имеющего запах лимона; в Нигерии он усиленно поедает Andropogon (Ohiagu, Wood, 1976; Wood, 1978). T. trinervius поедает 9 видов злаковых, но не трогает
растущие рядом бобовые; он срезает мертвые растения, даже если они еще зеленые, но в отличие от некоторых других видов не делает запасов (Bodot,
- . Вероятно, наиболее специализированы в отношении питания термиты, обитающие в индо-малайской области и собирающие лишайники с верхушек деревьев (Hospitalitermes) или же поедающие картон, из которого построены гнезда других термитов. Хотя термиты в общем вегетарианцы, все они поедают избыточных или раненых собратьев по гнезду, возвращая таким образом компоненты их тканей в общий круговорот веществ.
Другая особенность питания, неоднократно возникавшая в процессе эволюции термитов,-это поедание органического вещества (гумуса), содержащегося в почве. Это свойственно трем подсемействам Termitidae: Termitinae, Apicotermitinae и Nasutitermitinae. К числу бактериальных симбионтов Termitinae, питающихся гумусом, принадлежат спирохеты и свободноживущие актиномицеты. Нити последних поддерживаются гибкими кутикулярными шипами, имеющимися в задней кишке (рис. 2.3), а в средней кишке они обитают в жидкости, заполняющей пространство за пределами перитрофической мембраны, где обычно находятся патогенные микроорганизмы (Bignell et al., 1981). Актиномицеты, живущие в почве, секретируют фенолоксидазы, которые разрушают древесинные компоненты гумуса, и возможно, что кишечные актиномицеты делают то же самое. Благодаря этим симбионтам эффективность переработки пищи у термитов достигает 54-93%, т. е. очень высока по сравнению с 27% у тараканов, 30-40% у двупарноногих многоножек, 33% у равноногих раков или 14% у клещей (Wood, 1978).
Еще по теме Термиты как разрушители растительных остатков:
- Интенсивность разложения растительных остатков
- Поступление растительных остатков и биологическая активность почв
- Глава 5. ТРАНСФОРМАЦИЯ РАСТИТЕЛЬНЫХ ОСТАТКОВ И ОРГАНИЧЕСКИХ УДОБРЕНИЙ В ПОЧВЕ
- Изменение состава растительных остатков в процессе их разложения
- РАЗЛОЖЕНИЕ РАСТИТЕЛЬНЫХ ОСТАТКОВ И ФОРМИРОВАНИЕ ПОДСТИЛКИ
- ЭКОЛОГИЧЕСКИЙ МЕТОД ПРИ ИЗУЧЕНИИ . РАСПАДА РАСТИТЕЛЬНЫХ ОСТАТКОВ
- В. Я. Частухин РАСПАД РАСТИТЕЛЬНЫХ ОСТАТКОВ В ЕЛОВЫХ ЛЕСАХ
- В. Я. Частухин РАСПАД РАСТИТЕЛЬНЫХ ОСТАТКОВ В СОСНОВЫХ БОРАХ
- ВЫВЕДЕНИЕ ЭЛЕМЕНТОВ ИЗ РАСТИТЕЛЬНЫХ ОСТАТКОВ ПРИ ВОЗДЕЙСТВИИ РАЗЛИЧНЫХ ГРУПП САПРОТРОФОВ
- Глава II ОБЩЕЕ ОБСЛЕДОВАНИЕ МИКОФЛОРЫ РАСТИТЕЛЬНЫХ ОСТАТКОВ В ЕЛОВЫХ ЛЕСАХ
- Животный и растительный мир как единое целое
- РАСТЕНИЕ И РАСТИТЕЛЬНЫЙ ПОКРОВ КАК ОПТИЧЕСКАЯ СИСТЕМА
- Растительный покров болот как индикатор генезиса, строения и глубины торфяных отложений
- СПИСОК РАСТИТЕЛЬНЫХ ВИДОВ, ИСПОЛЬЗУЕМЫХ КАК КОРМА ДЛЯ ДОМАШНЕЙ ПТИЦЫ В УСЛОВИЯХ ТЕПЛОГО И УМЕРЕННОГО КЛИМАТА
- ГУМАННЫЙ РАЗРУШИТЕЛЬ