ПРАВИЛА О. ГЕРТВИГА И ИХ ПРИМЕНЕНИЕ К АНАЛИЗУ РАЗВИТИЯ АМФИБИЙ, РЫБ, ПТИЦ И НАСЕКОМЫХ
Первое правило Гертвига: ядро занимает центр активной
протоплазмы (полностью или почти полностью свободной от желтка). В алецитальных и изолецитальных яйцах ядро расположено ближе к центру (рис. 41,Л), а у телолецитальных оно располагается эксцентрично, в анимальном полушарии.
Второе правило Гертвига: веретено деления ядра располагается в направлении наибольшей протяженности чистой протоплазмы (рис. 41,Б).
Ярким примером правильности этих утверждений служит
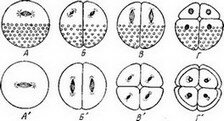
характер дробления телолецитальных яиц амфибий. Ядро у них, как уже сказано, расположено в анимальном полушарии. Наибольшая протяженность чистой протоплазмы параллельно экваториальной плоскости (см. рис. 42, Л и Л'). В этом направлении и окажется веретено первого деления ядра. Кариокинетический
 процесс связан с передвижением различных составных частей клетки, ее хромосом, частей цитоплазмы, кортикального слоя. Классическая описательная гистология и цитология, так много давшие биологии с - их методиками фиксации и окрашивания клеток, прививают, однако, изучающему клетки, ткани и развитие организма совершенно
процесс связан с передвижением различных составных частей клетки, ее хромосом, частей цитоплазмы, кортикального слоя. Классическая описательная гистология и цитология, так много давшие биологии с - их методиками фиксации и окрашивания клеток, прививают, однако, изучающему клетки, ткани и развитие организма совершенноне соответствующее действительности
представление о клетках, как о чем-то неподвижном. Мы неоднократно будем убеждаться в том, что такое представление о клетках необоснованно.
Различного рода закономерным движениям составных частей ядра и цито-
плазмы могут мешать зерна и пластинки желтка. Если желток расположен очень компактно, то процесс дробления в определенном участке яйца может подавляться.
Первая борозда дробления у яиц амфибий определяется указанным направлением веретена деления ядра (параллельно экваториальной плоскости, иначе — перпендикулярно к анимально- вегетативной оси яйца). Поэтому дробление произойдет в плоскости одного из меридианов яйца (рис. 42, А и А').
Положение веретен 2-го дробления и расположение новой борозды дробления также подтверждают правила Гертвига: в каждом из двух бластомеров веретена дробления располагаются, как и при 1-м дроблении, параллельно экваториальной плоскости, под прямым углом к бывшему направлению веретена 1-го дробления. Это происходит потому, что наибольшая протяженность чистой протоплазмы остается параллельной экватору и плоскости 1-го дробления. В результате произойдет снова меридиональное дробление. Борозды 1-го и 2-го дроблений появляются сначала на анимальном полюсе или вблизи от. него и постепенно врезаются в вегетативное, перегруженное желтком полушарие (рис. 42, Б и Б'). Итак, первые два дробления яиц амфибий — полные и равномерные. Однако после двух дроблений наибольшая протяженность чистой протоплазмы оказывается в направлении анимально-вегетативной оси, и именно в этом направлении расположены веретена третьего деления ядер во всех образовавшихся четырех бластомерах. Это направление перпендикулярно к экваториальной плоскости и плоскостям первых дроблений, в связи с чем очередное дробление окажется экваториальным (рис. 42, Г и Г'). Дробление опять-таки происходит полное, но уже
* иол°жение веретен дробления сбоку и с анимального полюса:
ложеннё "бл астомеппв еЦпг л Б’ Б' ~ второе дробление, В, В'- третье дробление; Г, Г'- по- лиженне оластомеров после третьего дробления
9S
.неравномерное; третья борозда дробления разделяет каждый бластомер на неодинаковые половины — верхняя (анимальная) меньшего размера, а нижняя (вегетативная), содержащая желток, большего размера. Затем две меридиональные борозды при четвертом дроблении сегментируют яйцо на 16 бластомеров, а две экваториальные борозды пятого дробления дробят яйцо на 32 бластомера. В дальнейшем меридиональные и экваториальные борозды чередуются, что отчасти также подтверждает правила Гертвига, но теперь уже вступают в действие не менее мощные факторы: взаимное давление бластомеров, процессы изменения желтка, различные явления взаимозависимости частей развивающегося эмбриона. Дробление перестает быть синхронным, и, как уже говорилось, в одних участках зародыша темп деления
 может усилиться, а в других— замедлиться; появляются и клетки, идущие по пути дегенерации.
может усилиться, а в других— замедлиться; появляются и клетки, идущие по пути дегенерации.
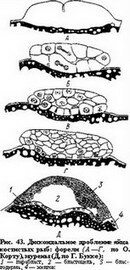
Не менее показательны другие примеры, подтверждающие правила Гертвига: развитие птиц (см. рис. 32), костистых рыб (рис. 43), а также рептилий. У яиц костистых рыб первое, второе, третье, а нередко и четвертое дробления меридиональные. Каждый из образовавшихся 16 бластомеров остается соединенным с желтком, полного обособления клеточных территорий не происходит. Зародышевый диск имеет незначительный размер по сравнению с массой желтка. При пятом дроблении яиц костистых рыб четыре центральных бластомера делятся экваториально, вследствие чего один из каждой образующейся пары бла- стомеров отделяется от желтка и приобретает вид клетки с вполне обособленной кле-
точной территорией, а другой из этой пары остается под ним в протоплазме, соединенной с желтком (рис. 43, Б, В). В это время каждый из 12 периферических бластомеров делится меридионально, в направлении формирующегося бластодиска (бластодерма), причем один из каждой пары сыновних бластомеров оказывается «краевым» бластомером, связанным с желтком, а другой — свободным, лежащим центрально. Однако в одном участке бластодиска (будущая его задняя сторона) меридиональная борозда проходит параллельно поперечному диаметру бластодиска. Связанные с желтком периферические бластомеры образуют краевую зону бластодиска. В этой зоне самые крайние бластомеры в течение всего периода дробления сохраняют связь с желтком. По мере того как такие «синцитиальные» бластомеры в краевой зоне, а также и подобного характера бластомеры под всем бластодиском в результате очередных дроблений все более беднеют цитоплазмой, образование новых клеток постепенно прекращается, однако ядра размножаются. Совокупность этих ядер называют перибластом или парабластом, или также мероци- тами (рис. 43,Д,1). Они играют роль в переработке желтка. Постепенно формируется бластула, состоящая приблизительно из тысячи клеток, иногда с выраженной полостью, наполненной жидкостью (бластоцель). Бластула снизу ограничена парабластом.
Принципиально такую же картину в смысле применимости правил Гертвига к анализу закономерностей дробления мы видим и у птиц (см. рис. 32).
В бластуле птиц различают: наружный слой центральной
части бластодиска — это плотный слой мелких бластомеров; под ними, в полости с жидкостью находится большое количество бластомеров, также не соединенных с желтком; дно полости и ее баковые стенки состоят из крупных периферических бластомеров. Постепенно, без деления клеток, происходит некоторое изменение в строении бластулы, а именно: бластомеры полости бластулы подходят к поверхностному слою бластодиска, клетки приобретают вид эпителия, который надвигается и ложится на периферическое поле бластодиска. Центральная часть зародыша благодаря просвечивающей жидкости полости бластулы оказывается более светлой; она называется area pellucida — светлое поле, а периферическая часть бластодиска, лежащего на желтке,— area ораса — темное поле.
Как видим, правила Гертвига вполне применимы к развитию телолецитальных яиц. Некоторые эмбриологи (П. П. Иванов, 1945) считают, что и дробление центролецитальных яиц не противоречит этим правилам,хотя объяснение характера поверхностного дробления центролецитальных яиц, исходя из принципов Гертвига, не может быть сделано с такой же ясностью, как это осуществлено в отношении телолецитальных яиц.
О влиянии распределения желтка на характер дробления можно судить и на основании экспериментов. Так, центрифугированием только что оплодотворенных яиц лягушки можно добиться еще большего уплотнения желтка в вегетативном полушарии, в результате чего яйцо начинает дробиться дискоидально.
Ниже показана зависимость характера дробления от того или иного типа яиц.
Схема зависимости формы дробления от типа яиц
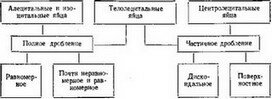
Итак, все факты в совокупности позволяют установить связь между типами дробления яиц, их количеством и распределением в них желтка, хотя биохимический и биофизический механизм этой связи в сущности до сих пор неизвестен. Эмбриология может говорить лишь о том, что желток как «инертный материал» является механическим препятствием при дроблении. На самом деле, как увидим далее, вопрос этот гораздо сложнее.
Источник: Токин Б. П.. Общая эмбриология: Учеб, для биол. спец, ун-тов.—4-е изд., перераб. и доп. 1987
Еще по теме ПРАВИЛА О. ГЕРТВИГА И ИХ ПРИМЕНЕНИЕ К АНАЛИЗУ РАЗВИТИЯ АМФИБИЙ, РЫБ, ПТИЦ И НАСЕКОМЫХ:
- Правила борьбы с болезнями и применения различных препаратов
- Применение органического анализа для изучения химических процессов в организме
- РАЗМНОЖЕНИЕ И РАЗВИТИЕ РЫБ
- ПРАВИЛА БЕЗОПАСНОСТИ ПРИ РАБОТЕ С РЕАКТИВАМИ, ПРАВИЛА ИХ ХРАНЕНИЯ
- Приметы для прогноза погоды по поведению насекомых, птиц и животных
- Развитие насекомых
- Влияние температуры на развитие насекомых.
- Амфибии
- Земноводные, амфибии
- ЦИЛИАТОЗЫ РЫБ Хилоденеллез рыб
- «Правила» эволюции групп
- 13.1.5. Эмпирические правила эволюции групп
- Глава IV ОБЩИЕ ПРАВИЛА ВЫРАЩИВАНИЯ БРОМЕЛИЕВЫХ
- Основные правила биосистемного проектирован ия
- ОСНОВНЫЕ ПРАВИЛА УСТАНОВКИ И ЭКСПЛУАТАЦИИ ПРИБОРОВ
- Правила приготовления компоста и перетоя
- 7.2. ПРАВИЛА ПЕРЕВОЗКИ ЖИВОТНЫХ РАЗНЫМИ ВИДАМИ ТРАНСПОРТА
- ОСНОВНЫЕ ПРАВИЛА СБОРА, СУШКИ И ХРАНЕНИЯ РАСТИТЕЛЬНОГО ЛЕКАРСТВЕННОГО СЫРЬЯ