СЛУЧАИ, ПРОТИВОРЕЧАЩИЕ ПРАВИЛАМ О. ГЕРТВИГА
Изучая эмбриональное развитие паразитических перепончатокрылых, О. М. Иванова в 1954 г. обнаружила, что у наездников в связи с паразитическим образом жизни происходит уменьшение размеров яиц и обеднение их желтком. Изменяется и тип дробления. Вместо поверхностного, сбойственного насекомым, дробление становится настоящим тотальным (полным), например у Copido- soma gelechiae. Имеются и случаи, не поддающиеся объяснению. Яйца Presiwichia aquatica, Car aphr actus redact us и Aphidius fabarum совершенно лишены желтка. Несмотря на это, дробление яиц первых двух' форм типичное поверхностное от начала до конца.
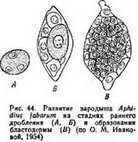
Своеобразно дробление яйца Aphidius fabarum. Это очень маленькие яйца, всего 86X30 мкм. Несмотря на отсутствие желтка, дробление сначала, как и обычно для насекомых, неполное, деление ядер не сопровождается разделением цитоплазмы (рис. 44^4), но через 5 ч после начала дробления происходит обособление клеточных границ, яйцо сразу распадается на
несколько (до 10) бластомеров (рйс. 44,5). О. М. Иванова *(1954) полагает, что поверхностный тип дробления насекомых, возникший в связи с накоплением желтка и закрепленный наследственно, продолжает действовать еще некоторое время и после исчезновения желтка. Эти суждения справедливы, если предполагать, что эволюция форм дроблений происходила не только в связи с изменениями в количестве и распределении в яйцах желтка, но и в связи с изменением каких-то других неизвестных механизмов.
 Накопилось много парадоксальных фактов, требующих создания новых гипотез. Так, например, установлено, что летние яйца ракообразного Moina paradoxa беднее желтком, чем зимние, но дробление их частичное, а богатые желтком зимние дробятся по голобластическому типу.
Накопилось много парадоксальных фактов, требующих создания новых гипотез. Так, например, установлено, что летние яйца ракообразного Moina paradoxa беднее желтком, чем зимние, но дробление их частичное, а богатые желтком зимние дробятся по голобластическому типу.Большой интерес представляют начатые А. Г. Гурвичем в 1908 г. опыты по центрифугированию яиц. Можно подвергнуть яйца морского ежа Arbacia punctulata вскоре после оплодотворения такому центрифугированию (в течение 2—5 мин), что ядро разрушится, и ядерный материал можно обнаружить лишь в виде глыбок, окрашивающихся по Фёльгену. Яйцо сохраняет сферическую форму, через несколько минут после центрифугирования начинается дробление при полном отсутствии каких-либо следов, веретена дробления. В небольшом проценте случаев такие яйцеклетки развивались даже до хорошо плавающих личинок — плу- теусов. Результаты опытов по центрифугированию трудно поддаются анализу.
Следует отметить, что А. Г. Гурвич и Л. Я. Бляхер объясняют возможность дробления яиц после энергичного центрифугирования не тем, что не разрушается гипотетический каркас. Их предположения основаны на представлении о природе протоплазмы, состоящей из неравновесных молекулярных констелляций (см. гл. XIII).
Известны явления, которые необъяснимы с точки зрения правил Гертвига. Таковы, например, результаты опытов Гарвей по партеногенетической активации лишенной ядра части цитоплазмы яйца морского ежа, в результате которой все же происходит дробление.
Многие другие факты также требуют особого объяснения; к ним, строго говоря, неприменимы правила Гертвига. Так, например, равномерное дробление свойственно, как правило, яйцам с равномерным распределением желтка, однако изолецитальные яйца некоторых млекопитающих дробятся неравномерно. Вероятно, очень важную роль при всех типах дробления играют еще недостаточно изученные типы контактов между бластомерами, характер межбластомерных пространств, большая или меньшая поверхность соприкосновения между бластомерами, явления взаимного скольжения клеток, образование на поверхности клеток псевдоподий, выростов протоплазмы, возможность амебоидных движений и иные явления. Неподвижность бластомеров — лишь кажущееся явление. Вспомним, например, яйца нематоды. В развитии этих яиц наблюдается перемещение бластомера Р2 и EMSt и образование ромбовидной фигуры на стадии четырех бластомеров (см. рис. 8). Активные перемещения бластомеров характерны и для спирального типа дробления. Причинный механизм этих явлений остается пока невыясненным.
Источник: Токин Б. П.. Общая эмбриология: Учеб, для биол. спец, ун-тов.—4-е изд., перераб. и доп. 1987
Еще по теме СЛУЧАИ, ПРОТИВОРЕЧАЩИЕ ПРАВИЛАМ О. ГЕРТВИГА:
- Нарушение правил транспортировки животных.
- Нарушение правил по уборке и утилизации трупов животных.
- Случаи с гостя ми-охотника ми и браконьерами
- Известно немало случаев возвращения лошадей...
- Сьюэлл Райт, адаптивные ландшафты и случай
- Центры круглосуточной неотложной ПОМОЩИ в случае отравления
- Директор лесного хозяйства без охотничьего билета — случаи с охотниками и охотвластями
- Выставка собак
- СУДЕБНАЯ ВЕТЕРИНАРНО-САНИТАРНАЯ ЭКСПЕРТИЗА ПРОДУКТОВ ПИТАНИЯ И СЫРЬЯ ЖИВОТНОГО ПРОИСХОЖДЕНИЯ
- ОЦЕНКА КАЧЕСТВА КОРМОВОЙ МУКИ ЖИВОТНОГО ПРОИСХОЖДЕНИЯ
- могильникAquila heliaca Sav.