Континуализм и структурализм
Среди множества этих принципиальных вопросов один — безусловно основной: есть ли вообще в природе экосистемы, или это понятие, придуманное для удобства? Множество ответов на этот вопрос можно расположить вдоль градиента, на концах которого расположены две альтернативные концепции экосистемы.
Распределение точек зрения по этому градиенту не равномерно, а резко бимодально, что позволяет говорить о существовании в экологии двух парадигм: континуализма (редукционизма) и структурализма (см., например, Миркин и др., 2002). В англоязычной литературе они получили, соответственно, названия индивидуалистическая гипотеза (individualistic concept) и гипотеза сообществ-единиц (community units).Парадигма континуализма была впервые сформулирована советским геоботаником Л.Г. Раменским (1884-1953) (Раменский, 1924) и независимо от него американским геоботаником Henry Allan Gleason (1882-1975) (Gleason, 1926). В данный момент она является абсолютно доминирующей как в нашей стране, так и за рубежом. С позиций континуализма написаны практически все учебники экологии и биогеографии. Согласно этой парадигме, как её понимают наиболее последовательные сторонники, виды эволюционируют более или менее независимо, воздействия видов друг на друга в сообществе, как правило,
«Виды встречаются вместе не потому, что приспособились друг к другу, а потому, что приспособились к общей среде» (Миркин, 1985: 51).
неспецифичны. Виды образуют сочетания случайным образом, в зависимости от локальных абиотических условий. Эти случайные сочетания видов — экосистемы, биоценозы, биогеоценозы, сообщества и т. п. — в природе плавно переходят одно в другое, образуя континуум, заполняющий неоднородную среду, как вода в луже заполняет неровности на поверхности земли. Некоторые исследователи даже считают континуальность основным свойством биосферы[*******]. Если всё же обнаруживают более или менее резкие границы сообществ, то их всегда считают созданными не биотой, а внешними факторами, например,
«Трудно отрицать наличие полной взаимонезависи- мости отдельных видов растений даже внутри сообщества...
последней классификационной единицей растительных сообществ являются не ассоциации, а виды растений» (Вальтер, 1968: 12-13).резкими физико-географическим градиентами. Пример: классический объект, рассматриваемый в качестве экосистемы, — озеро. Фактически это означает признание только двух уровней организации жизни: организм (вид, популяция) и биосфера и отрицание существования каких-либо самоорганизующихся биологических структур уровня, промежуточного между видом и биосферой. Континуализм подразумевает, что ответ биоты всегда одинаков на одинаковые изменения абиотических условий.
Альтернативная парадигма была впервые сформулирована американским геоботаником Frederic Edward Clements (1874-1945), в 1910-х гг. он работал в университете штата Миннесота, в 1917-1941 гг. руководил экологическими исследованиями в Институте Карнеги в Вашингтоне, ввёл понятия «сукцессия», «климакс», «биом», «экотон». В 1970-80-х гг. парадигма была существенно переработана и развита советским геоботаником С.М. Разумовским (1929-1983) и его последователями. Она не имеет общепринятого названия. Согласно этой парадигме, биосфера обладает эндогенной структурой, поэтому мне кажется наиболее правильным назвать её структурализм[†††††††]. Дискретные структуры, из которых, согласно структурализму, состоит биосфера, имеют отчётливые границы, проявляют функциональную активность, направленную на поддержание своей целостности, и эволюционируют как единое целое. Виды в процессе эволюции не приспосабливаются к некоей «внешней среде», а становятся элементом экосистемы. Ответ экосистемы на внешние воздействия часто зависит от видового состава и стадии сукцессии, т. е. может быть весьма различным.
Сторонники структурализма не только обосновывают свою точку зрения, но и объясняют, откуда берётся континуум в работах континуалистов. К сожалению, конструктивного диалога между сторонниками этих двух парадигм не происходит. Критика континуалистов сводится, в лучшем случае, к отрицанию отдельных фактов и положений структуралистов, всегда в самом мелком пространственно-временном масштабе, где различия между ними не столь очевидны и не всегда существенны.
Обычно же о существовании противоположной точки зрения даже не упоминают, а если и упоминают, то нередко с существенными её искажениями, приписыванием ей положений и высказываний, на ошибочность которых структуралисты сами прямо указывали. Так, например, в недавно вышедшей книге Б.М. Миркина с соавт. (2002), в таблице 13 сравнивают представления о сукцессии континуалистов и структуралистов по 6 пунктам. При этом сравнивают современные представления континуалистов и взгляды основоположника структуралистов- Clements (начало прошлого века). Если же сравнивать современные представления и тех, и других, то три из шести приписываемых в этой таблице структуралистам представлений прямо объявил неверными ещё четверть века назад С.М. Разумовский (1981).
Нечёткость понятий «экосистема», «биоценоз», «биогеоценоз», «сообщество» вполне приемлемы в рамках парадигмы континуализма, поскольку задачи исследований и не требуют этой чёткости (ибо априорно отвергают возможность существования самоорганизующихся единиц биосферы). В рамках же структурализма, признающего наличие таких структур, чёткость определений существенна.
Обе концепции существуют с начала века, пользуясь переменной популярностью, в зависимости от господствующей моды и научной школы, определяемых не в последнюю очередь объектом исследования и используемыми методами. И континуалисты, и структуралисты основывают свою позицию на фактических данных, полученных в результате исследования природных сообществ.
Континуализм сформирован в значительной степени западноевропейскими и североамериканскими геоботаниками, особенно второй половины XX в. В связи с этим надо отметить, что в Европе, по-видимому, уже много веков отсутствуют ненарушенные, т. е. естественные, не изменённые вмешательством человека сообщества, в Северной Америке они почти повсеместно исчезли в XX в. О том, как они выглядели, можно строить более или менее правдоподобные гипотезы. Диапазон этих гипотез весьма широк.
На одном краю находится гипотеза, согласно которой лесные массивы, не подвергавшиеся сплошной рубке несколько сотен лет (т. е. меньше максимальной продолжительности жизни видов-эдификаторов), являются естественными ненарушенными сообществами.
На другом краю находится гипотеза, согласно которой такие леса необходимо рассматривать как антропогенные промышленные древостой, стоящие гораздо ближе по своим свойствам к сельскохозяйственным полям, и, уж безусловно, они не являются естественными ненарушенными сообществами. Более того, всё большее признание получает точка зрения, что и сами пояса бореальных (тайга) и неморальных (широколиственные) лесов, а также степи и тундры, и многие пустыни имеют антропогенное происхождение. Согласно этой точке зрения, эти пояса возникли на месте экосистем, эдификаторами которых были крупные травоядные (мамонты и другие хоботные, носороги, парно- и непарнокопытные), экосистемы эти имели мозаичный характер, в них чередовались лесные и безлесные участки (Восточно-европейские леса..., 2004).
Структурализм сформирован североамериканскими, советскими и скандинавскими геоботаниками, изучавшими лесные сообщества, хотя и сильно нарушенные, но всё же в значительно меньшей степени, что и позволило им обнаружить эндогенную структуру биосферы.
«Сама методика случайной выборки, основанная на убеждении об отсутствии в природе объективных границ, никогда не сможет их выявить. Эта методика по своей сути заранее предопределяет результаты исследования» (Киселева, Разумовский, Расницын, 1969: 130).
В науке всегда построение моделей предшествует сбору информации. Иными словами, то, какой парадигмы придерживается исследователь, в значительной степени определяет его подход к изучению проблемы, методику отбора материала и его анализ, а, следовательно, и результаты. Так, широко применяемая при отборе проб практика игнорирования дискретности, часто заметной невооружённым глазом (закладка случайно расположенных площадок, не- соотнесение размера площадок с мозаичностью, т.
е., обычно, слишком крупный их размер и т. д.), приводит к тому, что существенная часть площадок захватывает площади, занятые несколькими ассоциациями видов. Математическая обработка такого неадекватно собранного материала с неизбежностью даёт картину континуума, независимо от действительной ситуации. В изучении бентоса случайный, т. е. часто нерепрезентативный характер расположения проб — существенная проблема, которую до некоторой степени компенсирует слабая мозаичность самих донных сообществ на больших глубинах.Выбираемая методология соответствует априорно предполагаемой природе исследуемого объекта. Не удивительно, что фактические материалы, собранные и обработанные континуалистами в соответствии с их представлением о биосфере как континууме, «объективно доказывают» существование континуума. Удивительным было бы иное. Не удивительно и то, что изучение тех же самых сообществ структуралистами даёт прямо противоположный результат.
В качестве примера рассмотрим интерпретацию данных по распределению видов континуалистами (рис. 4.1) и структуралистами (рис. 4.2). Первый рисунок (рис. 4.1) с незначительными вариациями (иногда дают только один рисунок или, наоборот, несколько) широко воспроизводят при обсуждении различий двух парадигм как экспериментальное подтверждение наличия континуума и отсутствия дискретных сообществ (см. например, рисунки в: Одум, 1975: 189; Уиттекер, 1980: 123 и 125; Бигон и др., 1989: 124-125; Brown, Lomolino, 1998: 105; Петров, 2001: 113; Воронов и др., 2002: 97; Сох, Moore, 2003: 52, 73; Нинбург, 2005: 6 и др.). Сравнение его с рис. 4.2 демонстрирует различия в подходах, осмыслении и интерпретации результатов. Главное отличие этих рисунков — не распределение кривых по оси абсцисс и их форма, как видится континуалистам, а то, что отложено по оси абсцисс. Континуалисты анализируют распределение видов по градиенту фактора, который они считают значимым, без какой-либо привязки к местности. Такой подход логичен при изучении экологических амплитуд отдельных видов и уже выделенных сообществ.
Однако реальные сообщества, распределены не только в воображаемом гиперпространстве факторов, но и в обыкновенном, вполне реальном географическом пространстве. Более того, сообщество (экосистема и т. п.) — это прежде всего комплекс видов, занимающий место в реальном, географическом пространстве. Пространство факторов — виртуальное, и в нём распределены виртуальные группировки организмов, которые тоже называют сообществами. Свойства реального и виртуального пространств различны, основное их различие в том, что в виртуальном пространстве факторов возможно на-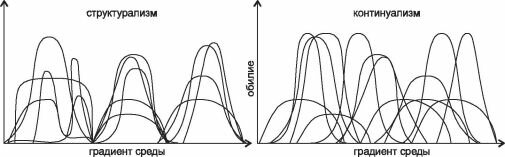
Рис. 4.1. Различия структурализма и континуализма в представлении континуалистов (каждому
виду соответствует своя кривая). 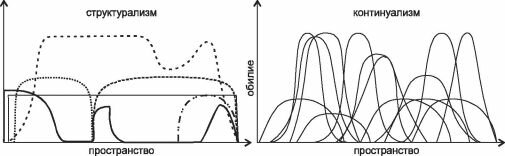
Рис. 4.2. Различия структурализма (разные кривые — разные виды) и континуализма (каждому виду соответствует своя колоколовидная кривая) в представлении структуралистов.
хождения в одной точке любого числа видов и сообществ, в географическом же — только одного. Происходит, таким образом, подмена понятий, и рисунок, призванный иллюстрировать различия взглядов представителей разных парадигм на распределение видов, в действительности иллюстрирует различия в подходах к сбору и анализу данных, т. е. вовсе не относится к обсуждаемому вопросу — к фактическому распределению видов по поверхности Земли. Графики, подобные изображённому на рис. 4.1, показывают распределение видов и их групп по градиенту изучаемых факторов, но никоим образом не проливают свет на вопрос о степени интегрированности реальных сообществ на местности. Более того, методы, используемые континуалистами для выделения сообществ (классификация и ординация) с точки зрения структуралистов в действительности вовсе не являются методами выделения экосистем, а лишь методами выявления экологических групп видов, сходно реагирующих на факторы внешней среды, поскольку выделение этих «сообществ» происходит не в географическом пространстве, а в «пространстве» факторов, что вовсе не одно и то же.
Эта идея многим покажется крамольной и абсолютно неприемлемой, поскольку общепринято считать наоборот. Поэтому давайте изучим этим методом (случайных проб и анализа их в пространстве факторов) территорию, несомненно разделённую в пространстве резкими границами на участки. Например, территорию Европы, поделённую на страны. На территории каждой из стран преобладают её подданные, поэтому будем считать территорию, на которой преобладают подданные одной страны, частью этой страны. Попробуем провести границы стран методами континуализма. Для этого подсчитаем число подданных разных стран в случайно расположенных квадратах, размером, например 50 х 50 миль, использованных для биогеографического районирования Северной Америки (Hagmeier, Stults, 1964). Очевидно, что часть этих квадратов попадёт на границы и захватит несколько стран. Уже только это снизит резкость границ. Рассмотрим далее распределение подданных разных стран в пространстве любых факторов, например, таких несомненно важных для экономики и населения этих стран факторов как увлажнение и температура. Даже и без проведения расчётов, совершенно очевидно, что мы не сможем получить чётких границ между странами, вместо этого мы получим континуум. А между тем границы эти существуют и необычайно резки (не имеют ширины в изучаемом масштабе). Задача разграничения государств вполне аналогична задаче разграничения сообществ, поскольку и те, и другие занимают какую-то часть поверхности Земли. Становится очевидным, что выбранные факторы не связаны с существованием дискретности. Если выбрать существенные для структуры факторы (экономические или социальные показатели, долю населения с тем или иным гражданством) чёткая дискретность скорее всего будет обнаружена. Проблема заключается в том, что исследователь изначально считающий, что дискретности нет, после первого анализа (по увлажнению и температуре) объявит, что математический анализ данных свидетельствует об отсутствии дискретности и наличии континуума, между тем, как математически доказана только отсутствие их связи. Сколь бы чёткими ни были границы, анализ несущественных факторов неизбежно даст нам в результате не чёткие границы, а плавный континуум. Выявить существенные факторы (которыми могут быть и биотические взаимоотношения) удаётся далеко не всегда. Например, один из процветающих отрядов насекомых — перепончатокрылые — наполовину состоит из паразитов. Однако не известно ни одного вида перепончатокрылых, паразитирующих на позвоночных, хотя есть паразиты, паразитирующие на паразитах позвоночных.
«Наблюдения подтверждают принцип Раменского и Глизона о существовании континуума сообществ: широкие перекрытия экологических амплитуд и рассредоточенность центров распределения популяций вдоль градиента среды приводят к тому, что большинство сообществ непрерывно переходят одно в другое, и это случается гораздо чаще, чем образование определённых ясно разграниченных друг от друга сообществ» (Уиттекер, 1980: 125).
Причина этого неизвестна, но факты это не меняет. Таким образом, невыявление дискретности с помощью таких методов не может служить доказательством отсутствия чётких границ. Появление континуума в результате такого анализа вовсе не отменит реальность границ, что легко проверить на пункте паспортно-визового контроля, попытавшись, вместо предъявления визы, рассказать пограничникам теорию о континууме в гиперпространстве факторов.
«Факты искажают отображение действительности в нашем сознании, если идея, их группирующая, порочна» (Фёдоров, 1987, цит. по 2004: 340).
Надо также помнить о почти повсеместном уничтожении или существенном преобразовании естественных сообществ. Безусловно, трудно провести резкую границу между лесом и степью, особенно, если степь распахали, а лес вырубили, но это не аргумент в пользу того, что эта граница и в естественных условиях (до вмешательства человека) — неясная и расплывчатая, а лес плавно переходит в степь, образуя экотон, —лесостепь. Таким образом, мнение континуалистов о том, что виды в нормальных условиях образуют не взаимно адаптированные комплексы, а континуум, на котором разные виды в зависимости от факторов внешней среды образуют разнообразные случайные сочетания (см., например, Уиттекер, 1980; Миркин, 1984; Миркин, Наумова, 1997 и др.), основанное на изучении распределения видов по градиентам факторов среды, не только не выдерживает никакой критики, но выглядит крайне наивно и неожиданно. Тем не менее, представление это парадоксальным образом почти общепринято считается доказанным. Однако число людей, придерживающихся какой-либо точки зрения, не является аргументом в поисках истины.
Только непосредственное изучение распределения видов по поверхности Земли (в географическом пространстве, а не в пространстве факторов) способно выявить реально существующие сообщества, хотя и здесь есть свои сложности. В частности необходим учёт сукцессионной динамики. Однако континуалисты и структуралисты вкладывают разный смысл в понятие «сукцессия» (см. ниже), поэтому и учитывать сукцессионную динамику они будут по-разному.
Появление понятия континуума и, особенно, его широкое распространение в геоботанике и, отчасти, в гидробиологии (Миркин, Розенберг, 1979; Миркин, 1987; Несис, 1977; Погребов, 1982; 1988), в значительной мере обусловлено не только недостаточным учётом сукцессионной динамики (а зачастую — и полным её игнорированием), но и широким распространением в современной экологии математических методов, не учитывающих пространственное распределение видов. Справедливости ради надо сказать, что существуют модификации широко распространённых методов, с помощью которых можно учесть реальное расположение мест отбора проб в пространстве, но они не входят в наиболее распространённые пакеты статистики. Исследователи, следуя традициям (моде?) не ставят перед собой цель учесть пространственное распределение изучаемых явлений, более того, булыпая часть биологов об их существовании и не подозревает. Иными словами, устойчивость представлений континуалистов обусловлена в значительной степени априорным игнорированием пространственной компоненты биологических (в данном случае — экологических) взаимодействий.
В последние десятилетия, не в силах отрицать очевидное, т. е. существенное влияние на распространение видов биологических факторов: конкуренции, эдификаторной роли и т. п., концепция континуализма претерпела некоторые изменения. Об этих изменениях Б.М. Миркин писал: «Суть новых взглядов на континуум заключается в том, что независимость распределений видов более не абсолютизируется, произошла «фитоценологи- зация континуума» (Миркин, Наумова, 1998). Виды в растительных сообществах занимают не фундаментальные, а реализованные экологические ниши, объём которых регулируется конкуренцией. Влияют на распределения видов и неконкурентные средообразующие взаимоотношения, например, влияние эдификаторов (таких, как ель в бореальных лесах или сфагнум на верховых болотах). По этой причине распределения видов по градиентам среды могут быть не только симметричными колоколовидными, но и асимметричными и даже би- и полимодальными» (Миркин, 2005: 523).
«На одной из своих лекций Давид Гильберт[‡‡‡‡‡‡‡] сказал:
— Каждый человек имеет некоторый определённый горизонт. Когда он сужается и становится бесконечно малым, то превращается в точку. Тогда человек говорит: «Это моя точка зрения» (Физики шутят, 1993: 44).
Как ясно из приведённой цитаты, нового в действительности очень мало: по-прежнему происходит подмена понятий. Виды влияют на распределение друг друга на местности, в реальном пространстве, а не по «градиентам среды», не в воображаемом пространстве экологических факторов. Что здесь: непонимание сущности спора или нежелание изменить свою точку зрения?
Введение пространства в классические законы экологии меняет их нетривиальным образом.
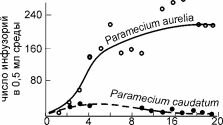
Так, согласно одному из них — принципу конкурентного исключения — «два вида, занимающие одну и ту же экологическую нишу, не могут устойчиво сосуществовать, поскольку в результате конкуренции один из видов будет вытеснен другим» (Гиляров, 1990: 159). Итальянский математик Вито Вольтерра показал это на системе дифференциальных уравнений. Выпускник кафедры зоологии беспозвоночных МГУ Г.Ф. Гаузе на опытах с инфузориями в небольших стеклянных сосудах, получил экспериментальное подтверждение расчётов Вито Вольтерра. В опыте, результаты которого приведены на рис. 4.3, один из видов вымер через 20 суток после начала опыта (что примерно соответствует 40 генерациям). Позднее аналогичные результаты были получены на многочисленных опытах, и этот закон стали считать одним из основных законов экологии, названным по имени первооткрывателей законом (принципом) Вольтерра-Гаузе. Изучение же ситуации в природе показывает, что сосуществование видов с идентичными потребностями (нишами) — явление достаточно обычное. Например, фитопланктон по всему Мировому океану и в пресных водах состоит из большого числа видов одноклеточных водорослей с очень сходными нишами, причём виды эти встречаются вместе и, по-видимому, сосуществуют многие тысячелетия (если не миллионы лет), но до сих пор друг друга не вытеснили. Этот факт никак не соответствует теории Вольтер- ра-Гаузе, но тем не менее до сих пор не получил общепринятого объяснения, а лишь название «планктонный парадокс» (Hutchinson, 1961). Аналогично, число видов деревьев, совместно произрастающих в тропическом лесу, очень велико, в то же время различия их ниш показать не удаётся. Подобные различия теории и фактов пытаются объяснить двумя способами.
время, сут
Рис. 4.3. Динамика популяций инфузорий Paramecium aurelia и P. caudatum в совместной культуре По G.F. Gause, 1934 из: Гилярова (1990).
- Предположением о неполноте фактов:
- поскольку согласно дифференциальным уравнениям Вито Вольтерра достаточно различия в одном измерении ниши, а число измеряемых исследователем в природе параметров всегда конечно, предполагают, что на самом деле виды имеют различные ниши, просто исследователи эти различия не обнаруживают.
- Введением дополнительных условий, при соблюдении которых виды с идентичными потребностями всё-таки могут сосуществовать, например:
- неоднородности среды: в опытах Г.Ф. Гаузе, в которых в среду вводили неоднородности (например укрытия), виды с близкой биологией сосуществовали на протяжении всего эксперимента;
- наличие плотностно-зависимых эффектов: ограничение численности разных видов не межвидовыми (конкуренцией за ресурсы, например), а внутривидовыми взаимодействиями. Анализ уравнений (Chesson, 2000; Adler et al., 2007) показывает, что имеются значения коэффициентов, при которых возможно сосуществование видов с идентичными требованиями (нишами).
В приведённых выше моделях события развиваются во времени. Предложенные для их объяснения теории также объясняют развитие процесса во времени, хотя реальные события развиваются не только во времени, но и в пространстве. Экспериментальные сосуды Г.Ф. Гаузе с точки зрения инфузорий (т. е. учитывая их способность к перемещению в пространстве) равноценны одной точке.
Между тем, существует множество работ, в которых показано, что учёт пространства может существенно изменить результаты. Проиллюстрируем это наглядной моделью, основанной на идеологии клеточных автоматов[§§§§§§§] (Грабовский, 2005). В ней особи двух или нескольких видов (число видов можно задать от двух до десяти) взаимодействуют на совершенно однородном моделируемом поле. Все особи всех видов имеют равную конкурентоспособность и занимают одну нишу (имеют идентичные потребности). С помощью модели можно изучить динамику во времени: (1) суммарной по полю относительной численности видов и (2) распределение отдельных видов в пространстве. Пространство измеряют в размерах особей, время — в генерациях. Измерения этих параметров в относительных, а не абсолютных величинах (метры, сутки и т. п.) биологически более правильно (Азовский, 2003). При размножении каждая особь делится на несколько эквивалентных ей потомков (можно задать разное число потомков). Каждый потомок расселяется в пределах некоторого задаваемого радиуса. Если выбранный участок оказывается свободным — занимает его. Если участок занят другим видом, то расселяющаяся особь с вероятностью 0,5 (виды имеют равную конкурентоспобность!) либо вытесняет его, либо вымирает. При половом размножении добавляется ещё один задаваемый параметр: радиус поиска полового партнёра (разноса гамет, пыльцы, спермиев, неоплодотворённых яйцеклеток и т. п.). Модель показывает, что виды с одинаковой конкурентоспособностью, занимающие одну нишу в однородном пространстве, могут сосуществовать весьма долго, если это пространство достаточно велико. Изменение параметров модели (радиуса расселения, радиуса поиска полового партнёра, размера поля, числа потомков) влияет на результат. На модели получено, что продолжительность сосуществования видов уменьшается при:
- уменьшении отношения размера экспериментального поля к радиусу перемещения особей (т. е. по мере приближения поля к точке), предельный вариант (поле равно точке) с максимальной неустойчивостью сосуществования видов и рассматривает теория Га- узе-Вольтерра;
- увеличении плодовитости.
«Формулировку закона Ома необходимо уточнить следующим образом: «Если использовать тщательно отобранные и безупречно подготовленные исходные материалы, то при наличии некоторого навыка из них можно сконструировать электрическую цепь, для которой измерения отношения тока к напряжению, даже если они производятся в течение ограниченного времени, дают значения, которые после введения соответствующих поправок оказываются равными постоянной величине» (Физики шутят, 1993: 144).
«Условия экспериментов, породившие «принцип Гаузе», не имитируют природную ситуацию, а наоборот, исследуют поведение и судьбу популяций, в экстремальных... ситуациях, которых нет в природе» (Фёдоров, 1987, пит. но 2004: 362).
При низкой способности к расселению и большом радиусе поиска полового партнёра (ситуация типичная для, например, деревьев тропических лесов, имеющих крупные плоды, которые разносятся на расстояние гораздо меньшее, чем пыльца), большое число видов (в данной модели максимальное число — 10) с идентичными нишами способны существовать весьма долго. Прогон модели с параметрами фитопланктона (радиус расселения мал, размножение бесполое) даёт ещё более устойчивую ситуацию сосуществования 10 видов с полностью идентичными экологическими нишами. Виды образуют пятна, форма пятен довольно изменчива, положение пятен более постоянно. Получаемые на моделируемом поле пятна разных видов весьма напоминают наблюдения в природе. Такая картина получается и при изучении распределения сообществ (примерно соответствующему распределению наиболее массовых видов) инфузорий на литорали Белого моря (Бурков- ский, 1992, 2006: 154) (рис. 4.5). По результатам съёмок с интервалом в 2 недели (примерно 30 поколений) форма пятен сильно изменилась, но многие пятна остались примерно в тех же местах. Таким образом, учёт пространства существенно изменяет результаты. Более того, клеточные автоматы позволяют смоделировать ситуации, для которых не удаётся подобрать систему уравнений и коэффициенты. При этом получаемые результаты вполне соответствуют наблюдениям в природе не только по моделируемым свойствам, но даже и по тем, которые вовсе никак не учитывали при построении модели. Кроме уже разобранных свойств одновидовых пятен, вполне соответствует наблюдениям в природе и распределение относительных частот видов по результатам модели (рис. 4.4а). Таким образом, оказывается, что противоречие между фактическими данными и обычно использованными математическими моделями, в частности системой дифференциальных уравнений Воль- терра, объясняется, по-видимому, непригодностью выбранного математического аппарата для моделирования таких процессов. Аналогично результаты эксперимента Г.Ф. Гаузе можно экстраполировать на природную ситуацию только в тех условиях, когда соблюдаются сделанные при его постановке допущения (особи способны расселяться на расстояние, сопоставимое с размерами однородного участка, в котором происходит взаи-

Рис. 4.46. Распределение видов в пространстве в том же опыте.
Начальное распределение видов случайное, на рисунке приведено распределение через 100 поколений. Оттенки серого соответствуют тем же видам, что и на рис. 4.4а.

Рис. 4.4а. Относительная численность десяти видов с идентичными нишами в однородном пространстве через 100 поколений. Результаты одного из прогонов модели при следующих параметрах: размер экспериментального поля 75 х 75, радиус расселения особей 1, радиус поиска полового партнёра 10, плодовитость 4 потомка, разные оттенки серого соответствуют разным видам). Время эксперимента (100 поколений) примерно в 2,5 раза больше, чем в опыте Г.Ф. Гаузе на рис. 4.3.
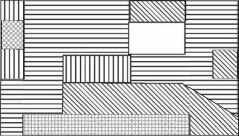
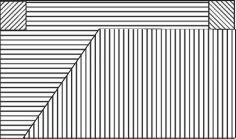
мейобентос
макробентос
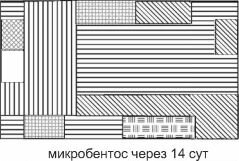
микробентос
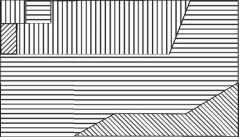
Рис. 4.5. Структурная гетерогенность сообщества на уровне макро-, мейо- и микробентоса.
Тип штриховки соответствует определенной структуре доминирования в микро-, мейо- и макробентосе, выявленной кластерным анализом. Исследованная площадь — 3200 м2, 25 станций (по: Бурковский, 1992). С увеличением размеров организмов возрастает и абсолютный радиус их расселения (в метрах), возрастание абсолютных размеров и радиуса расселения приводит к уменьшению абсолютной мозаичности одновидовых пятен. Динамика положения пятен микробентоса хорошо соответствует прогнозируемым моделью В.И. Грабовского.
модействие видов). Построенная на основании этих моделей и экспериментов теория нуждается в существенной уточнении, поскольку в основе её лежат артефакты. В частности, закон Гаузе-Вольтерра — «два вида с одинаковой биологией не могут сосуществовать в однородной среде» — следует дополнить словами «в одной точке», поскольку в пространстве, а тем более в природе, они сосуществовать вполне могут, о чём свидетельствуют и наблюдения.
Еще по теме Континуализм и структурализм:
- 6-11. Диасеть и структурализм
- Моделирование континуальности и дискретности
- Изменение таксономического разнообразия по поверхности Земли
- Ценотическая система Ценотическая система — что это такое?
- Методы группирования регионов
- О свойствах климакса
- Изменение биологии видов в разных частях ареала
- Причины и скорость образования новых видов и экосистем
- Кружево ареала
- 10-4. Родство и сущность