Изменение таксономического разнообразия по поверхности Земли
Очевидное различие биоразнообразия в разных частях Земли стимулировало поиски закономерностей, описывающих этот феномен. На западе выявление этих закономерностей — одна из основных задач макроэкологии.
Были предложены закономерности, описывающие постепенное снижение видового богатства организмов в направлении от тропиков к полюсам, с увеличением глубины или высоты местности, связь размеров ареалов с широтой местности и т. д. и т. п. При анализе таких закономерностей, прежде всего, возникает проблема того, какие регионы следует сравнивать. Очевидно, что сравнивать следует однотипные регионы, но что считать однотипным сильно зависит от используемых концепций. С позиций континуализма, отрицающего эндогенную дискретность биосферы, вполне допустимо сравнение списков любого произвольно выбранного региона (но всё же желательно примерно одного размера, чтобы уменьшить влияние размеров площади). С позиций структурализма следует сравнивать лишь биогеографические регионы одного ранга. Поскольку размер биогеографических регионов весьма слабо связан с их рангом, сравнение списков регионов, равных по площади, но разного биогеографического ранга скорее будет отражать различие рангов, нежели изменение биоразнообразия вдоль анализируемого градиента. Так, часто пишут о большем таксономическом разнообразии шельфовой биоты по сравнению с глубоководной. При этом оперируют данными, полученными в результате сравнения числа видов на разных глубинах или на разных широтах, обычно для всего Мирового океана. Между тем всё океаническое ложе занято единым биогеографическим выделом, вероятно, ранга области. На шельфе же расположено несколько областей. Не удивительно, что в одной области обитает видов меньше, чем в нескольких. То же справедливо и в отношении широтных различий: шельф севера Северного полушария занят атланто-аркто-тихоокеанской областью, тогда как в тропических районах областей как минимум две. Даже при одинаковом числе видов на область, очевидно, что, суммируя данные по нескольким областям, мы неизбежно получим большую цифру, чем для одной области.А.И. Азовский (Hillebrand, Azovsky, 2001; Азовский, 2003) проанализировал результаты 153 исследований и показал, что «убывание числа видов от экватора к полюсам действительно является устойчивой, статистически достоверной тенденцией видового разнообразия для самых разных систематических групп... В то же время сила градиента закономерно зависит от характерных размеров тела особей: чем крупнее организмы, тем сильнее выражен у них широтный градиент. Так, число видов деревьев, млекопитающих и пойкилотермных позвоночных обнаруживает, как правило, сильную отрицательную корреляцию с широтой, тогда как для зоопланктона, мейофауны и одноклеточных (как простейших, так и микроводорослей) подобная корреляция выражена очень слабо или отсутствует вовсе» (Азовский, 2003: 174).
Реликты и рефугиумы
Особой категорией видов являются реликты (англ. relict от лат. relinquere — оставить позади). Следует различать филогенетические и биогеографические реликты. О филогенетических реликтах говорят в случае таксонов ранга, обычно, не ниже семейства, представленного в данный момент лишь одним (или немногими), но ранее многочисленными видами. Например, Latimeria является единственным современным представителем ранее весьма разнообразной группы кистепёрых рыб, а несколько видов неопилин — немногими современными представителями ранее богатого видами класса моллюсков Мо- noplacophora. Филогенетические реликты ещё называют живыми ископаемыми.
О биогеографических реликтах говорят в случае, если данная местность ранее входила в сплошной ареал таксона (ЦС), а ныне изолирована от его основного ареала.
В 1947 г. филогенетическим и биогеографическим реликтам была посвящена специальная сессия французского биогеографического общества. На ней не только было рекомендовано различать эти понятия, но и предложено использовать для них разные термины.
Термин «реликт» (relicte, англ. эквивалент — relict) было предложено использовать только для биогеографических реликтов, а филогенетические реликты называть relique (англ. эквивалент — relic). Рекомендациям этим, однако, не следуют.Термин реликт применяют и к надвидовым системам (сообществам, экосистемам). Реликтовые сообщества представляют собой изолированные части ЦС, которые ранее имели в этом районе сплошной ареал, но при изменении климата или по какой-либо иной причине изменили своё положение на Земле (или вымерли). Виды, из которых состоят реликтовые сообщества, если они не входят в состав окружающей ЦС, естественно, будут биогеографическими реликтами. Поскольку сообщества состоят из многих видов, районы, занятые реликтовыми сообществами являются районами, в которых сконцентрировано большое число биогеографических реликтов. Такие районы получили название рефу- гиум (англ. refuge, лат. refugium, мн. число — refugia). Если данные ЦС и виды по-прежнему где-то существуют, а их нахождение в данном месте является следствием более широкого ареала в прошлом, то для таких изолированных участков ареала используют также термин эксклав. Часто биогеографические реликты являются и филогенетическими.
Число обитающих в рефугиуме видов по определению меньше числа видов во всей ЦС (иначе это был бы не рефугиум). Из-за отсутствия соответствующих видов некоторые сукцессионные стадии (ассоциации) невозможны. Поэтому сукцессии в рефугиумах часто завершаются диаспорическими субклимаксами. Устойчивость рефугиумов тем выше, чем более полно представлены в нём сукцессионные ряды, т. е. чем больше в нём ассоциаций. Число ассоциаций проще всего оценить (с определённой ошибкой) по числу видов: чем больше в рефугиуме видов, тем больше в нём ассоциаций. Для устойчивости ЦС наиболее важны блоки, организованные по консортному типу — блоки низших трофических уровней. Блоки высших трофических уровней до определённой степени могут быть заменены другими и тем легче, чем больше в них пастбищных черт.
Центры разнообразия и центры происхождения
Рис. 5.35. Центры разнообразия жуков рода скрытоглавы азиатские. По Г.М. Абдурахманову и др. (2001).
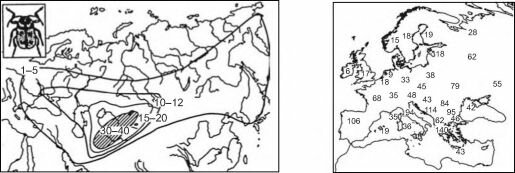
Рис. 5.36. Центры разнообразия покрытосеменных рода смолёвка Silene. По Thompson, 1973).
Широкое принятие «араратской» концепции объясняет многочисленные попытки выяснить место происхождения таксона надвидового ранга путём картирования числа видов (или других подчинённых таксонов). Для этого на карту наносят число таксонов[§§§§§§§§§§§], отмеченных в каком-либо достаточно маленьком районе и проводят изолинии (пример на рис. 5.35). При этом выявляются места, где видов много, и места, где их мало. Сам факт неравномерного распределения числа видов внутри ареала изучаемого таксона надвидового ранга сомнению не подлежит. Но при интерпретации необходимо помнить, что такие карты обычно построены по сравнительно небольшому числу точек, поэтому более или менее достоверными на них являются положение пиков и общий тренд. Границу ареала изучаемого таксона, как обычно, проводят с изрядной долей экстраполяции. Что касается положения изолиний на таких картах, то их проводят с гораздо большими экстра- и интерполяциями на основе недоказанного предположения о плавном изменении числа видов в пространстве, т. е. эти линии — не факты, а их интерпретация. Гораздо более корректным было бы нанесение самих данных на карту, но это делают гораздо реже, возможно потому, что картинки получаются не столь красивые (рис. 5.36).
Часто центры таксономического разнообразия интерпретируют как центры происхождения таксона, из которых таксон постепенно расселяется радиально. Такая интерпретация в неявном виде подразумевает два допущения:
- Периферические виды отличаются по своему возрасту от видов, обитающих в центре (дисперсионистская биогеография).
- Виды в центре происхождения образуются симпатрически, причём этот способ видообразования резко доминирует в изучаемом таксоне.
Относительно первого допущения имеются две диаметрально противоположные точки зрения.
Согласно одной виды, обитающие в центре происхождения, древнее таксонов, обитающих на периферии. Такого мнения придерживаются основатель кладиетики энтомолог Hennig и другой энтомолог (Brundin). Согласно другой точке зрения, которой, в частности, придерживается такой известный биогеограф (тоже энтомолог), как P.J. Darlington (1966), новообразованные виды вытесняют предковые на периферию ареала, так же считает и J.C. Briggs (1974, 1992, 1995, 2003, 2004). Эта точка зрения согласуется с данными по распределению возраста герматипных кораллов (рис. 3.59), хотя герма- типные кораллы и не естественный таксон, а жизненная форма, да к тому же, это не единственное возможное объяснение уменьшения возраста таксонов из центра разнообразия к периферическим районам. Второе допущение, вроде бы, не очень согласуется с представлением континуалистов о преимущественном происхождении видов путём географической изоляции, хотя, с точки зрения структуралистов, это вполне возможно.Итак, само по себе наличие повышенного числа видов рода в каком-либо месте может свидетельствовать, а может и не свидетельствовать о месте происхождения данного рода. В любом случае, для суждений о месте происхождения таксона необходимо привлечение дополнительных данных, желательно палеонтологических. Локальное повышение числа видов таксона может лишь привлечь внимание исследователя для последующей интерпретации, не более.
Для перемещения ареала таксона или сообщества в пространстве в геологическом масштабе времени используют термин «прохорез».
Использование термина «центр происхождения» не только подчеркивает факт повышенного числа таксонов в данном месте (что легко определить), но и утверждает, что они и произошли здесь (а это доказать гораздо сложнее). Поэтому места повышенного таксономического разнообразия лучше называть не центрами происхождения, а центрами разнообразия. Часто, особенно в группах, по которым палеонтологические данные скудны или вовсе отсутствуют, приходится строить гипотезы о месте происхождения таксона на основе анализа распространения его современных представителей.
При этом надо учитывать, что высокое таксономическое разнообразие ЦС (группы ЦС) может быть обусловлено разными причинами:- Большим потоком энергии через её ареал. В этом случае таксономическое разнообразие свойственно многим таксонам.
- Большим размером её ареала. В этом случае таксономическое разнообразие свойственно многим таксонам.
- Сильной её фрагментацией. В этом случае таксономическое разнообразие свойственно таксонам с низкой способностью к расселению.
- Сильной её изоляцией. В этом случае вспышку таксономического разнообразия дают только некоторые таксоны, при этом многие таксоны, свойственные аналогичным ЦС, отсутствуют.
- Данный регион — рефугиум. Определить это можно по другим группам с хорошими палеонтологическими данными.
А.Н. Миронов (1983, 1985) выдвинул гипотезу, согласно которой повышенное число видов наблюдают не только в центрах происхождения, где выше скорость видообразования, но и в центрах аккумуляции, где более эффективно аккумулируются виды, произошедшие где-то в другом месте. Для характеристики способности района к аккумуляции видов А.Н. Миронов ввёл понятие «таксономической ёмкости района». Реальность центров аккумуляции он показал на нескольких примерах, два из которых приведены ниже.
- Анализ фаунистических связей на видовом уровне показывает значительное родство батиальных фаун Индо-Вестпацифики и Карибского бассейна. Многие виды, обитающие в этих районах различимы с трудом. Причём большинство видов, распространённых только в этих районах, принадлежат к родам, неизвестным в ископаемом состоянии (по-видимому, они возникли совсем недавно). Причина такого сходства, вероятно, — недавнее формирование батиальной биоты Карибского бассейна. Существовавшая ранее здесь батиальная биота в значительной степени вымерла во время плейстоценовых оледенений из-за понижения продуктивности поверхностных вод (Prell, Hays, 1976) и резкого изменения режима осадконакопления (Emiliani et al., 1975; Be et al., 1976). Факт массового вымирания тропической фауны Западной Атлантики в это время показан для моллюсков (Stanley, Campbell, 1981). Последующее изменение условий в этом районе потребовало формирования новых ЦС, основным источником видов для которых были районы со сходными условиями, а также местные биоты. Индо-вестпацифическая батиальная биота, вероятно, и ранее была наиболее разнообразной, поэтому её вклад в новые ЦС оказался, в итоге, наиболее существенным. Кроме Карибского бассейна, существенный вклад индо- вестпацифическая батиальная биота сделала и в биоты некоторых других недавно сформировавшихся районов: расположенного в восточной части Тихого океана хребта Наека (Парии и др., 1997) и западноатлантических гор Метеор (Миронов, Крылова, 2006).
- В миоцене возрос температурный градиент между экватором и полюсами и установилась циркуляция полярного типа и, следовательно, увеличилась продуктивность планктона Северной Пацифики. Одновременно произошло значительное увеличение площади Северной Пацифики (возникли Охотское и Берингово моря). Всё это привело к возникновению обширной абиссальной территории с новыми, ранее не существовавшими в таком масштабе условиями — с богатыми органикой глубоководными илами. На этой территории сформировалась своя ЦС (или несколько). Наиболее преадаптированными к условиям абиссали Северной Пацифики были виды, населявшие антарктическую абиссаль, где аналогичные изменения произошли раньше (циркумантарктическое течение возникло ещё в конце палеогена). Это обусловило наблюдающееся сейчас высокое сходство таксономического состава абиссали Антарктики и Северной Пацифики.
Аккумуляция таксонов при формировании новой ЦС — нормальная и даже обязательная стадия. Вероятно, ещё одним центром аккумуляции была бореальная Западная Атлантика : многие амфибореальные виды, проникшие в Атлантику из Пацифики не смогли закрепиться в Восточной Атлантике (хотя ряд известен в ископаемом виде), но сохранились в гораздо более таксономически бедной Западной Атлантике.
Еще по теме Изменение таксономического разнообразия по поверхности Земли:
- Таксономический состав и разнообразие макробентоса
- Площадь листовой поверхности посевов кукурузы
- ТЕМПЕРАТУРА ПОВЕРХНОСТИ ЯЧЕЕК С РАЗВИВАЮЩИМИСЯ ЧЛЕНАМИ ПЧЕЛИНОЙ СЕМЬИ
- Изменение размеров животных с изменением температурных климатических условий.
- Таксономическая структура и современные подвиды '
- Таксономические категории и система классификации
- ТАКСОНОМИЧЕСКИЙ СОСТАВ ПОЧВЕННЫХ ПРОКАРИОТ
- ЭКОЛОГО-ТАКСОНОМИЧЕСКИЙ СОСТАВ ПОЧВЕННОЙ ФАУНЫ
- 9-1. Рост разнообразия и скоростей
- Пляска полюсов Земли
- Таинственная «дрожь земли