Изменение биологии видов в разных частях ареала
Возможно, О. Drude (1876) первый заметил, что, хотя экологические потребности вида примерно постоянны по всему ареалу, положение стаций с подходящими условиями меняется. Это закономерность была затем описана в виде многочисленных правил, или законов: правило предварения (Буш, 1917/1918; Алёхин, 1926, 1936), Raplaceability of Ecological Valence (Wamecke, 1936), Relative Habitata Constancy (Walter, Walter, 1953), правило смены ярусов (Гиляров, 1959), правило смены местообитаний (Бей-Биенко, 1966, 1971).
Все эти закономерности, законы и правила описывают происходящие изменение распределения видов и сообществ по стациям с изменением абиотических условий в био- географическом масштабе. Некоторые из этих закономерностей стали предметом изучения макроэкологии. С точки зрения структурализма в этом нет ничего удивительного, поскольку в этих случаях один и тот же вид или сходные физиономически ассоциации и сообщества, входят в разные ЦС, и поэтому занимают разные ниши.северная граница ареала
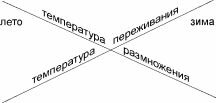
лето зима
южная граница ареала
Рис. 5.11. Правіша Ортона и Хатчинса. По А.Н. Голикову, О.А. Скарлато (1972).
В гидробиологии также были предложены частные варианты этой закономерности. Один из них касается размножения гидробионтов. Размножение вида происходит при примерно одинаковой температуре воды. У широко распространённых видов, обитающих в местах с большой амплитудой температуры (литораль и верхняя сублитораль) необходимая температура бывает в разные периоды года. Поэтому у видов, нерест которых происходит в период прогрева водоёма, в более тёплых частях ареала размножение происходит в более холодное время и наоборот. Было даже предложено правило — правило Уртона (J.H.
Orton), согласно которому минимальная температура размножения определяет северную границу ареала, а максимальная температура размножения — южную границу ареала.Для красоты это правило было дополнено также температурой выживания — правило Хатчинса (L.W. Hutchins): минимальная температура выживания определяет северную границу ареала, максимальная температура выживания — южную границу ареала (рис. 5.11).
Необходимо отметить, что сезонные колебания температуры характерны почти исключительно для поверхностной водной массе. Поэтому не удивительно, что данные по распространению в североевропейских морях нижнесублиторальных видов полихет (наши данные), ракообразных и иглокожих (Blaker, 1984) не подтверждают правило Хатчинса.
Частое отсутствие связи распространения видов с температурой обусловлено, по- видимому, тем, что температура влияет опосредованно. При значениях температуры, выходящих за рамки экологической амплитуды ЦС, в которую входит вид, одну ЦС обычно сменяет другая. Если вид не входит в эту другую ЦС, то скорее всего он не будет найден в её ареале, хотя температурные условия для него могут быть вполне приемлемыми. Например, условия в глубоководной части Северного моря соответствуют условиям склона Северной Атлантики, однако обитающая там ЦС не может попасть в глубоководную часть Северного моря, поскольку они разделены обширными мелководьями. Две ЦС могли бы заселить глубоководную часть Северного моря: шельфа Северного моря, но для неё глубина слишком велика, и глубоководная арктическая, но для неё слишком высока температура. В результате глубоководная часть Северного моря заселена модифицированной ЦС шельфа Северного моря, в которую вошли некоторые виды глубоководной арктической (Maldane arctica, Amage auricula), обитающие здесь при температуре, постоянно гораздо более высокой, чем в других частях ареала этих видов.
Относительно же первой части правила данных очень мало, и сказать, действительно ли это правило или отдельные факты, нельзя, тем более, что есть и прямо противоположные факты.
Так, имеются виды, размножающиеся в период охлаждения воды, у них зависимость сроков размножения от времени года обратная: в более тёплых районах нерест начинается позже, в более холодных — раньше. Среди рыб к первой группе относятся весенненерестующая сельдь, треска, сардины, анчоус, ставриды, камбалы Атлантики и Пацифики, скумбрия, мерлина, золотистый окунь Атлантики. Ко второй группе — салака, осенненерестующая сельдь, треска Атлантики (Беренбойм, 1980; Марти, 1980). Существуют также виды, у которых есть оба типа размножения. У таких видов на тепловодной и холодноводной границах ареалов наблюдается смыкание сроков нереста двух рас. У промысловых рыб это было показано у сельдей в Ла-Манше (южная граница), салаки Ботнического залива и трески залива Св. Лаврентия (северная граница) (Беренбойм, 1980).Изменение температуры воды может происходить не только по оси север - юг, но и запад - восток, а также по глубине. Поэтому в более тёплых районах виды часто обитают глубже, это явление получило название субмергенция.
Структура ареала обсуждалась во многих работах (см. например, Беклемишев, 1969; Udvardy, 1969; Городков, 1986), однако в них не различали ценофильные и ценофобные виды. Между тем структура их ареалов принципиально различна.
Еще по теме Изменение биологии видов в разных частях ареала:
- Питание лося в разных частях ареала
- Изменения экосистем. Сопряженная эволюция разных видов (коэволюция)
- ОБЕЗВРЕЖИВАНИЕ НАВОЗА РАЗНЫХ ВИДОВ ЖИВОТНЫХ
- Сравнение ритмов разных видов подвижности и активности
- Формы взаимоотношений между популяциями разных видов растений
- 6.1. ПОДГОТОВКА ПАСТБИЩ ДЛЯ РАЗНЫХ ВИДОВ И ГРУПП животных
- ПОЛОВЫЕ ЦИКЛЫ У ЖИВОТНЫХ РАЗНЫХ видов
- ПОЛОВЫЕ ЦИКЛЫ У ЖИВОТНЫХ РАЗНЫХ видов
- Фармакокинетика фенбендазола в организме животных разных видов (В. Вшуе1, 1980)
- ОСОБЕННОСТИ ТЕЧЕНИЯ РОДОВ И ПОСЛЕРОДОВОГО ПЕРИОДА У ЖИВОТНЫХ РАЗНЫХ ВИДОВ
- Глава I МИКРОФЛОРА СОСНОВОЙ ДРЕВЕСИНЫ НА РАЗНЫХ ФАЗАХ РАСПАДА.СУКЦЕССИЯ ВИДОВ ГРИБОВ И БАКТЕРИЙ