Периодизация онтогенеза
Онтогенез представляет собой полную последовательность всех этапов роста и развития растения от диаспоры до отмирания особи. По более развернутому определению Л.А.
Жуковой (1997;5), «онтогенез — это генетически обусловленная полная последовательность всех этапов развития одной особи или ряда поколений особи от зиготы или любой диаспоры до естественной смерти на завершающих этапах вследствие старения». Такой полный цикл роста и развития генета можно рассматривать как большой жизненный цикл. Онтогенетический цикл, свойственный отдельному рамету от его возникновения до отмирания, называют малым жизненным циклом. Время, затрачиваемое особью на прохождение отдельного онтогенетического состояния, по предложению Л.А. Жуковой (1995), называют онтохроном. Продолжительность онтохронов очень разная в зависимости от таксономического вида растения и условий произрастания.Онтогенез по своей природе является непрерывным процессом, но в связи с изменениями в ходе этого процесса качественных и количественных параметров особей растений еще с XVIII века началась разработка концепции его дискретного описания. Первоначально в качестве отдельных этапов онтогенеза выделялись фенофазы. Первые регистрации фенофаз проводились в Англии, но достаточно определенную дискретную шкалу фенофаз впервые предложил К. Линней. Позже в связи с потребностями сельскохозяйственной практики и уточнением оптимальных сроков внесения удобрений и пестицидов стали разрабатываться более детализированные фенологические шкалы, подразделяющие онтогенез культурных и сорных растений на 6, 12, а в современных шкалах и на 100 самостоятельных фаз (этапов). Эти шкалы предназначены главным образом для периодизации онтогенеза однолетних культурных растений и сорняков (Злобин, Прасол. 1993).
Принципиальна.і особенность фенологических и связанных с ними шкал состоит в том, что они ориентированы на видовой уровень организации растительного покрова.
При их применении регистрируется состояние, в котором находится основная масса особей данного вида.С иных позиций систему дискретного описания онтогенеза (главным образом для многолетних трав) разработал Т.А. Работ- нов (1950). Эта система позже была усовершенствована и распространена на растения других жизненных форм А.А. Урановым, Л. Б. Заугольновой, О.В. Смирновой и другими специалистами. Она характеризует биологический возраст, или онтогенетическое состояние особей растений в тот или иной период времени, а главное исходит из того, что разные особи в популяции находятся в разном онтогенетическом состоянии, и, следовательно, ориентирована на выявление неидентичности особей в популяции. В настоящее время, поданным Л. А. Жуковой (1997), на основании популяционно- онтогенетического подхода изучено уже около 400 видов растений. В четырех томах «Онтогенетического атласа лекарственных растений* дано описание онтогенеза 147 видов растений.
В жизненном цикле растений принято выделять четыре основных онтогенетических периода;
- Латентный период — состояние жизнеспособных семян.
- Предгенеративный период - охватывает состояние от всходов до взрослых вегетативных растений.
- Генеративный период - соответствует цветущим и плодоносящим особям.
- Постгенеративный период, на протяжении которого особи утрачивают способность активно расти и размножаться и постепенно отмирают.
В свою очередь каждый из этих онтогенетических периодов подразделяется на ряд онтогенетических состояний. Схема этого деления предсталена в табл. 6.2.
Для основных онтогенетических этапов онтогенеза ключевыми признаками чаще всего являются следующие:
- семена (sc) - это жизнеспособные кепроросшие семена или не- раскрывающиеся плоды в почве. В популяционных исследованиях эту возрастную группу выделяют редко, так как извлечение семян из почвы представляет собой трудоемкую процедуру, которая требует специального оборудования. Еще сложнее определение жизнеспособности извлеченных из почвы семян;
|
О нтогенетиче ски й период |
Онтогенетические состояния растений |
Условное обозначение |
|
Латентный |
Семена |
sm |
|
Пре дге не дативный |
Проростки |
Р |
|
Ювенильные |
! |
|
|
Имматурные |
іт |
|
|
Виргинильные |
V |
|
|
Г оперативный |
Молодые генеративные |
9л |
|
Средние генеративные |
9г |
|
|
Старые генеративные |
9з |
|
|
Лостгенерэтивн.ый |
Субсенильные |
SS |
|
Сенильные |
$ |
|
|
Полностью отмершие |
SC |
- проростки (р) - молодые растения, сохраняющие связь с семенем и имеющие первичный корешок, семядоли, вынесенные на поверхность, или первичный листок. У злаков характерным признаком этого состояния является наличие колеоптиля;
- ювенильные растения (у) - отличаются упрощенной морфологической структурой, в частности, имеют ювенильные листья; связь с семенем отсутствует; побег чаще всего еще не ветвится;
- имматурныерастения (іт) - по морфологической структуре являются переходными от ювенильных к виргннильным и отличаются наличием ветвления побега. Имматурные растения у злаков соответствуют началу фазы кущения;
- виргинильные растения (о) - обладают всеми морфологическими признаками, присущими взрослым особям, но не цветут и не плодоносят. У плотно- и рыхлокустовых злаков вирги- нильное состояние соответствует фазе кущения. У корневищных злаков виргинильные растения представлены системой парциальных кустов, соединенных коммуникационными корневищами;
генеративные растения (g0~g3) - отличаются наличием цветков и плодов. У многолетних поликарпических растений это возрастное состояние часто подразделяют на 3-4 подэтапа в зависимости от осуществленного количества актов генеративного размножения. У молодых генеративных растений (gj генеративные органы формируются первый раз в их онтогенезе, такие растения еще сохраняют активный рост вегетативных органов. У среднегенеративных растений (g2) генеративное состояние достигает своей максимальной выраженности, а у старых (g%), хотя и сохраняется генеративная функция, но она ослаблена, а в вегетативной сфере начинается отмирание отдельных структур. Скрытогенеративное онтогенетическое состояние ?0 выделяют только для отдельных видов растений. В первую очередь это злаки, где оно соответствует фазе трубкования, а также ряд видов растений, у которых генеративные почки закладываются еще с осени и находятся в скрытогенеративном состоянии до следующего вегетационного периода. Во всех таких случаях наличие скрытогененративного состояния должно проводиться путем анализа почек с использованием бинокулярного микроскопа. Поскольку скрытогенеративное онтогенетическое состояние может быть выделено у генеративных растений в состояниях glt g2 и gy то его регистрация выпадает из общего принципа подразделения растений на молодые, средние и старые генеративные;
- субсенильныс (ss) и сенильные (s) растения - отличаются преобладанием процессов отмирания, цветение и плодоношение отсутствуют или имеют рудиментарный характер;
- отмирающие растения (sc) - не имеют живых надземных побегов вообще.
Принадлежность к определенному возрастному состоянию определяется по степени выраженности определенных морфологических (например, степень расчлененности сложного листа) и физиологических (например, способность дать потомство) признаков. Таким образом фиксируется, прежде всего, биологический возраст особи. Очень важно включать в такой анализ только качественные признаки, так как разнообразные количественные признаки, отражающие размер растения и его частей свидетельствуют не о биологическом возрасте, а о жизненном состоянии растения. В практике популяционной ботаники, к сожалению, этот принцип иногда нарушается, что приводит к неадекватной оценке состояния и особей, и структуры популяций. Б.М. Миркин и Л.Г. Наумова (1998) подчеркивали, что всегда есть «немалый риск принять не старое, но угнетенное растение за старое и, напротив, достаточно старое, но процветающее - за более молодое*.
Для определения онтогенетических состояний растений в настоящее время разработаны специальные ключи и выпущены атласы изображений (например, «Диагнозы и ключи возрастных
состояний... *, 1980-1989, под редакцией Л. А. Жуковой «Онтогенетический атлас лекарственных растений*, 1997-2004). Опубликовано довольно много работ с описанием состояния растений на разных этапах их онтогенеза (выпуски «Биологической флоры Московской области*, Горнов, 2005 и многие др.). В качестве примера на рис. 6.6 приведены основные онтогенетические состояния донника лекарственного MelUotus officinalis.
Дробность шкалы онтогенетических состояний, как и критерии их установления, разумеется, зависят от объекта и целей исследования. В ряде случаев шкала может быть редуцирована без ущерба для качества получаемой информации. Имматурное состояние быстро преходящее, а у многих видов растений вообще не выражено, поэтому имматурные и виргинильные особи часто можно рассматривать как одну группу вегетативных растений.

Рис. 6.6. Основные онтогенетические состояния особей донника лекарственного Melilotus officinalis (по JI.A. Жуковой, 1995).
Онтогенетическое состояние особей: р - всходы; j - ювенильные; іт - имматурные; о - виргинильные; g. - молодые генеративные; gt~ среднегенеративные; g3 - старые генеративные; ss - субсенильные; а - сенильные растенияДеление генеративных особей на gv, g2 и g3 состояния имеет смысл только тогда, когда генеративное размножение одной и той же особи может осуществляться на протяжении нескольких лет. Если генеративная функция осуществляется особью всего один раз в жизни, то регистрируется только возрастное состояние g. Так, например, у малины ее раметы имеют четкий двухлетний цикл. Побеги первого года всегда виргинильные, второго - генеративные, и расчленять их на подфазы g2 и g3 - значит не понимать самого смысла популяционно-онтогенетического метода.
Особым способом подразделяется генеративное состояние у однолетних растений. Здесь наиболее целесообразно онтогенетическое состояние gx оценивать как бутонизацию, g2 - как полное цветение Hgj- как плодообразование.
Нечетко различаются у отдельных видов субсенильное и сенильное состояние.
У раметов клоновых растений некоторые авторы (Harper, 1978) считают целесообразным выделять только три этапа онтогенеза: ювенильный, репродуктивный и постредродуктивный. Многие зарубежные исследователи и для популяций растений, формирующихся из генетов, предпочитают использовать членение онтогенеза не более чем на 3-5 онтогенетических состояний. Для обобщенных оценок структуры популяций этот подход вполне правомерен и достаточен.
Пока что слабо разработана периодизация онтогенетических состояний древесных растений. Наиболее удачную шкалу периодизации онтогенетических состоянии этих растений приводят О.В. Смирнова и М.В. Бобровский (2001). Она имеет следующий вид:
- gtgt; 82- 83 ~ молодые, средневозрастные и старые генеративные деревья, которые приступили к семяношению и поддерживают его в течение нескольких лет или десятилетий;
- s - сенильные деревья, имеющие вторичную крону с листьям или хвоей ювенильного типа, семяношение отсутствует.
При общем прогрессивном характере этой шкалы она сохраняет некоторые недочеты. В частности, авторы связывают ювенильные деревья с травяно-кустарничковым ярусом, а имматур- ные - с ярусом кустарников, тогда как фактически подрост древесных пород, особенно в неблагоприятных условиях, может долго оставаться в ярусе трав и мелких кустарничков, и его стволик ветвится. В этой сфере необходима дальнейшая работа, хотя в целом у древесных растений спектр календарных возрастов особей чаще всего дает более ценную информацию о популяции, чем спектр онтогенетических состояний.
Изучение онтогенеза многолетних растений показало, что в некоторых случаях, особенно при эколого-фитоценотических стрессах, растение может временно задерживаться в любом онтогенетическом состоянии на более или менее длительный период времени. Тогда это состояние продолжается дольше, чем это наблюдается в типичных случаях. Особенно примечательно, что такие задержки могут наблюдаться у потенциально генеративных растений, когда они на тот или иной период времени (от года до нескольких лет) теряют возможность формировать генеративные органы, хотя и ее утрачивают потенциал генеративности. Т. А. Ра- ботнов (1975), а затем О-В. Смирнова и др. (1984) предложили такие особи называть квазисенилъными. Термин не очень удачен, так как подлинная сенильность в этом случае не развивается и признаки таких особей не всегда носят старческий характер, для которого обязательно преобладание процессов отмирания модулей организма. Это своего рода диапауза растений, резкий спад жизненного состояния, ведущий к снижению темпов развития, а не изменению возрастности. Такие растения при улучшении условий быстро «оправляются* и продолжают развитие. Поэтому многие специалисты выделяют в генеративном периоде растений особый этап - временно нецветущие растения. Его можно обозна чать как g . Это более корректная терминология.
В шкале онтогенетических состояний каждая фаза рассматривается как отдельный этап в росте и развитии особи, отличающийся морфологической структурой, фиэиолого-биохимическ нм
из
состоянием и особым типом взаимоотношений с эколого-фитоце- нотической средой. К прохождению каждого онтогенетического состояния растения аккумулируют определенную долю энергии, переходящую в органические вещества его тела, что дало возможность А. А. Уранову (1975,1977) вычислить эту долю количественно и представить в виде индексов возрастности, которые удобны для оценки возрастности популяции в целом. Используется формула:
I*. ’
где Д - возрастноеть популяции;
к, - численность особен определенной возрастной группы; т - коэффициент возрастности данной группы.
Коэффициенты возрастности приведены в табл. 6.3. Поскольку переход особей из одного онтогенетического состояния б другое идет на протяжении вегетационного периода, для оценки состояния растений в популяции очень важен выбор периода времени его регистрации. К сожалению, этому вопросу в популяционной литературе уделялось мало внимания. Разумеется,
Таблица 6.3. Периодизация полного онтогенеза растений (по А.А. Ура нову, 1975иЛ.А. Жуковой, 1997)
|
Период онтогенеза |
Возрастное состояние j |
Условное обозначение |
Индекс возраст- ности по Уранову |
|
1. Латентный |
1. Покоящееся семя |
зе |
0,0025 |
|
И. Пред генера |
2. Проросток |
Р |
0,0067 |
|
тивный |
3. Ювенильное |
І |
0,0180 |
|
4. Имматурное |
іт |
0,0474 |
|
|
5. В ирги пильное |
V |
0,1192 |
|
|
Ш. Генера |
6. Скрытогенеративное |
9с |
|
|
тивный |
7. Молодое генеративное |
9gt; |
0,2700 |
|
в. Зрелое генеративное |
9г |
0,5000 |
|
|
9, Старое генеративное |
9з |
0,7310 |
|
|
IV. Постгенера- |
10. Субсенильное |
as |
0,8808 |
|
тиеный |
11. Сенильное |
5 |
0,9529 |
|
12. Отмирающее растение |
SC |
0 9819 |
нельзя регистрировать генеративность растения в период до его естественного перехода в эту фазу или после ее завершения. Поэтому наиболее разумный период для регистрации онтогенетического состояния растения (если не вести за ним постоянных наблюдений в течение всего вегетационного периода) - это срок от начала бутонизации до завершения плодоношения. На этом отрезке времени удобно регистрировать и онтогенетическое состояние всех остальных особей популяции, которые не вступили в фазу гене- ративности.
6.5.2. Онтогенетические спектры популяций
Концепция дискретного описания онтогенеза Работнова - Уранова оказалась очень удобной для целей популяционных исследований. Онтогенетические состояния достаточно легко регистрируются, особи одного онтогенетического состояния объединяются В одну группу, а при обработке материла просто и быстро вычисляется соотношение (в процентах или в долях единицы) в популяции растений разного онтогенетического состояния.
Во избежании путаницы JI.B. Заугольнова (1986), а затем
В.В. КричфалушийиГ.М. Мезев-Кричфалуший(1994)предложи- ли называть соотношение особей разных возрастных состояний в популяции ее возрастным спектром, тогда как соотношение особей разного календарного возраста - возрастным составом- Это разумно, но часто только из контекста работы видно, какой тип состава популяции анализирует тот или иной автор. Более удачным, на наш взгляд, является онтогенетический спектр (состав) популяции.
На основе соотношения в популяции особей растений разных онтогенетических состояний Т.А. Работнов предложил подразделять популяции на три основные категории:
- инвазионные - с преобладанием предгенеративных растений;
- нормальные, в которых доля особей разного онтогенетического состояния приблизительно сбалансирована и преобладают генеративные растения;
- регрессивные, в которых преобладают постгенеративные растения.
Позже к трем основным категориям была добавлена категория ложноинвазионных популяций, представленных в сообществе только почвенным банком семян. Кроме того, могут быть выделены вегетативно-омоложенные онтогенетические спектры, свойственные популяциям, сформированным из молодых растений вегетативного происхождения. Сукцесеионными популяциями называют популяции, в которых возобновительный процесс преобладает над отмиранием растений. Представителями московской популяционно -демографической школы регрессионные популяции были подразделены на четыре варианта (В1 — /14) по соотношению сенильных, субсенильных и отмирающих растений. В дальнейшем число категорий популяций за счет переходных типов было увеличено до девяти, однако это не привело к получению более ценной геоботанической информации. Большое дробление основных типов спектров нецелесообразно с прагматической точки зрения.
Точного количественного критерия для установления категорий популяций не было предложено. В этой связи Б.М. Миркин и Л.Г. Наумова (2001) разработали удобный полуколичвственяый критерий для оценки категории популяции (табл. 6.4).
Инвазионные популяции характерны для серийных типов растительных сообществ, а также для случаев внедрения в сообщество новых видов, нормальные свойственны видам растений, которые длительное время устойчиво сохраняются свою позицию в фитоценозе, регрессивные соответствуют ситуациям постепенного выпадения вида из сообщества.
Онтогенетические спектры могут быть полночленными с наличием в них представителей всех онтогенетических состояний и неполночленными (фрагментированными), когда особи того или иного онтогенетического состояния в популяции отсутствуют, В случае, когда популяции находятся в равновесном состоянии и их
Таблица 6.4. Классификация популяций по характеру онтогенетического спектра (поБ.М. Маркину, Н.Г. Наумовой и А.И. Со- ломещ, 2001)
|
Онтогенетическое состояние особей |
Категория ценолопуляции |
||
|
инвазионная |
нормальная |
регрессивная |
|
|
П редгене рати вные |
+++ |
+ |
_ |
|
Генеративные |
+ |
+++ |
+ |
|
Пост ген ерат ивные |
- |
+ |
+ + + |
т
онтогенетический состав лишь слегка флюктуирует по годам, их называют дефинитивными.
Дополнительно полезно онтоге нетич ес к ие спектры популяций растений оценивать последующим их характерным особенностям:
- левосторонний спектр- отличается преобладанием дегенеративных особей, что отражает наличие благоприятных условий для возобновительного процесса, а так как этот процесс непостоянен по годам, то такие спектры очень динамичны. Левосторонние спектры свойственны также популяциям, только начинающим внедряться в данный фитоценоз. Характерны они и для некоторых древесных видов в лесных сообществах;
- центрированный спектр - отличается высокой долей генеративных особей. Он характерен для многолетников с простым онтогенезом, а также для случаев, когда возобновительный процесс слабо выражен, а период пребывания особей в сенильном состоянии короткий. Центрированные спектры свойственны популяциям с устойчивым статусом в данном сообществе;
- правосторонний спектр - отличается высокой долей в популяции сенильных растений. Он характерен для видов с длительным периодом старения при ослаблении возобновительного процесса. Правосторонние спектры обнаруживаются и у популяций, выпадающих из данного фитоценоза;
- бимодальный спектр - имеет два пика; обычно один в предге- неративной и другой в сенильной части спектра. Он свойственен видам растений с активным возобновлением и длительным сенильным периодом. Характерен для многих злаков и осок.
По мнению Р.В. Попадюк и др. (1999), популяции с левосторонними, центрированными и бимодальными онтогенетическими спектрами являются устойчивыми и могут длительное время реализовывать сменяемость поколений.
Погодичные флюктуации онтогенетических спектров чаще всего происходят за счет молодой части популяции. Если же популяции свойственно направленное изменение состава, плотности или численности особей, то ее, как было отмечено выше, называют сукцессионной. Сами по себе сукцессионные популяции не могут быть долговечными: они или переходят в категорию дефинитивных, или исчезают.
Благодаря авторитету и широкой разветвленности московской популяционной школы Работнова - Уранова онтогенетическая структура популяций изучена у более чем 250 видов растений разных жизненные форм и в разных почвенно-климатических условиях. Об особенностях онтогенетических спектров популяций разных видов растений существует огромная литература. Приводимый в ней материал тем не менее не всегда репрезентативен, поскольку часто строится на данных, полученных в течение одного-двух вегетационных периодов. В работах В.Г. Кияк и др. (1988), И. Царик и др. (2001) на основании данных многолетних наблюдений за популяциями прямо подчеркивается, что в связи с погодичными флюктуациями одноразовые и одногодичные характеристики онтогенетического состава популяций малоинформативны. Необходимы данные многолетних учетов. И.Т. Попонова (2006), располагая 20-летними наблюдениями за популяциями арабидопсиса Arabidopsis thaliana, пришла к выводу, что даже у этого однолетнего растения для выявления характерной онтогенетической структуры популяций необходим минимальный срок наблюдений в 5 лет. Очевидно, для многолетних трав он значительно больше.
Конкретные онтогенетические спектры могут в той или иной степени соответствовать основным их категориям или отличаться от них некоторыми деталями. Варьирование онтогенетических спектров популяций обусловлено факторами, в которых протекает онтогенез растений. Поэтому онтогенетические спектры популяций несут важную информацию о ходе процессов возобновления и отмирания особей, о темпах смены поколений в популяциях и, следовательно, позволяют оценивать и прогнозировать динамические процессы в фитоценозах, которые составляют данные популяции.
Одним из распространенных вариантов популяций, как было отмечено выше, являются ложноинвазионные. Они свойственны видам растений, способным создавать длительно сохраняющиеся почвенные банки семян, клубней или корневищ. В отдельные благоприятные для них годы происходят массовые вспышки численности всходов таких растений, но они быстро отмирают, не переходя во взрослое или тем более в генеративное состояние. В состоянии ложноинвазионных популяций искусственно поддерживаются популяции однолетних сорных растений в аг- рофитоценоаах, когда после дождей или после уборки урожая массово появляются всходы этих сорняков, но их быстро уничтожают культивацией посева или гербицидами. Для истощения почвенного запаса семян таких видов специально используются приемы, провоцирующие прорастание семян: поверхностные боронования, дискование и др.
Для интегральной характеристики онтогенетической структуры популяций предложены количественные индексы, разработанные Л.А. Жуковой (1987) и позже модифицированные К. В. Глотовым (1998). Они основаны на соотношениях между разными онтогенетическими когортами, составляющими популяцию. Более удачный вариант вычисления индексов был разработан И.Н. Коваленко (2004), в котором оценивается доля каждой онтогенетической когорты по отношению к общей численности популяции.
Индексы И.Н. Коваленко имеют следующий вид:
- Индекс возобновляемости:
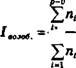
Формула
jh—10°-
100.
- Индекс генеративности:
![Формула]()
Формула
MHtp, Р-,
2gt;.
- Индекс старения популяций:
![Формула]()
Формула
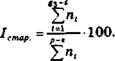
1-і
- Индекс общей возрастности популяции - вычисляется как отношение индекса старения к индексу возобновляемости данной популяции:
Эти индексы в совокупности позволяют достаточно точно охарактеризовать общее состояние популяции и особенно пригодны для сравнительного анализа популяций одного и того же вида растений при произрастании в разных эколого-ценотических условиях.
На основании сопоставления индексов возрастности популяций можно оценить скорость динамических смен в популяциях, используя формулу:
N -At
где V - скорость изменения возрастности популяции,
1 - индекс возрастности популяции в конце I2 и начале Р учет
ного срока соответственно;
N - число особей в популяции;
ДГ - продолжительность периода наблюдения.
Сравнение онтогенетических спектров двух популяций целесообразно осуществлять по методу, предложенному Л.А. Живо- товским (1979), с расчетом коэффициента сходства г и критерия идентичности популяций I:
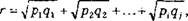
Формула
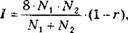
Формула
где Afj и N2 - объемы первой и второй выборок;
р, - частоты встречаемости того или иного признака в первой выборке;
Я, ~ частоты встречаемости того или иного признака во второй выборке;
8 - число этапов, на которые подразделен онтогенез.
При г = 1 онтогенетические спектры совпадают, при г = 0 полностью различны. Достоверность промежуточных значений находят по критерию идентичности по таблицам распределения с-. Если значения I превышают табличное значение, то онтогенетические спектры принимают статистически достоверно различными при принятом уровне значимости.
Большой фактический материал показывает, что устойчивое положение ценопопуляции в фитоценозе достигается при оптимальном равновесном сочетании в ней особей разного онтогенетического состояния. Для ценозообразующих видов растений в экологически стабильных условиях характерны долночленные дефинитивные онтогенетические спектры популяций с максимумом на генеративных особях, на долю которых может приходиться до 40- 70% их общей численности. Иногда они могут быть лево-, иногда правосторонними. Типичны нормальные онтогенетические спектры для растений степей (Боровик, 1994).
В разных фитоценозах онтогенетические спектры часто оказываются достаточно однообразными (Некратова, Некратов, 1987 и мн. др.). И.М. Ермакова (1987:27) верно подчеркивала, что «возрастные спектры - наиболее стабильные характеристики ценспо- пуляций *. Для большинства видов растений характерны нормальные популяции, иногда неполночленные со слабой бимодальностью. Например, такой оказалась структура популяций на высокогорных лугах Карпат (Кричфалуший, Комендар, 1990).
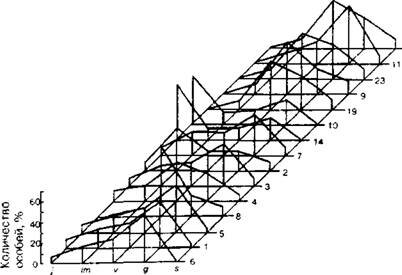
Рис. 6.7. Онтогенетические спектры Narcissus angustifolius в ) 5 разных типах фитоценозов в Карпатах. Типы фитоценозов обозначены арабскими цифрами (по В.В. Кричфалуший, 1988)
Наиболее подвижной их частью являлась группа проростков и ювенильных особей, доля которых в популяциях наиболее изменчива по годам (Климишин, 1987). Так, В.В. Кричфалуший (1988) исследовал онтогенетические спектры нарцисса узколистного Narcissus angustifolius в 15 разных типах растительных сообществ в Карпатах и, как видно из рис. 6.7, принципиальных различий между онтогенетическими спектрами не обнаружил, хотя в деталях по соотношению растений разных онтогенетических состояний спектры отличались. Анализ этих деталей часто дает очень важную экологическую и биологическую информацию.
При резкой смене эколого-фитоценотической обстановки в онтогенетических спектрах могут происходить существенные сдвиги. Например, у клевера лугового Trifoloium pratense на градиенте возрастания пастбищной нагрузки на луг популяции из нормальных с пиком на особях состояния g2 переходили в резко правосторонние с преобладанием старых генеративных и субсе- нильных особей (рис. 6,8).
Характер онтогенетического спектра зависит от вида растения с его спецификой вегетативного и генеративного размножения.

Рис. 6.8. Онтогенетическая структура популяций клевера лугового на трех участках пойменного луга с разным режимом использования (по Е.С. Кирильчук, 2007)
О.В. Смирнова и др. (2002) установили наличие определенного соответствия между жизненной формой многолетних трав и онтогенетическими спектрами их популяций. Для моноцентричес- ких многолетних трав наиболее характерны левосторонние спектры с преобладанием предгенеративных растений, для неявнопо- лицентрических трав - бимодальные с максимумами на предгенеративных и субсенильных растениях, а для явнополицентри- ческих - неполночленные с преобладанием генеративных растений. У поли центрических растений количество вегетативных партикул обычно больше, чем генеративных. Онтогенетические спектры редких и малообильных видов растений с небольшими и дизъюнктивными популяциями могут быть самыми разнообразными - от инвазионных до регрессивных. Двухвершинность онтогенетических спектров часто связана с антропогенными воздействиями на сообщество (Буланая, 1991).
Для лесных сообществ вусловиях климакса характерны левосторонние спектры с максимумом на растениях в онтогенетическом состоянии от проростков до виргинильных включительно. Такие спектры свойственны древесным растениям как при семенном, так и при вегетативном размножении (Смирнова и др., 1990).
Устойчивость популяций далеко не всегда непосредственно связана с их онтогенетическими спектрами. В ряде случаев в ценозах стабильно сохраняют свои позиции виды с неполночленными возрастными спектрами. Описаны и противоположные случаи. В.М. Остапко (2005), изучив онтогенетические спектры более чем 100 видов травянистых растений, в том числе многих редких на юго-востоке Украины, установил, что при низкой численности и малых размерах популяций для них остаются характерными спектры со стабильной онтогенетической структурой. На Алтае Gueldenstaedtia monophylla имеет дефинитивные популяции с центрированными спектрами, в которых 43-70% приходится на генеративные особи, но она является редким растением из-за низкой семенной продуктивности и высокой степени гибели всходов (Селютинаи др., 2008).
Для однолетних растений характерна сезонная сменяемость онтогенетических спектров: в начале сезона в популяциях преобладают предгенеративные растения, в середине - генеративные, а к осени - генеративные и сенильные. Порой онтогенетические спектры у многолетних трав по годам имеют волнообразно-сукцессион - ные изменения. Интересный случай волнообразно-сукцессионной смены популяций пальчатокореиника балтийского Dactylorhiza longifoiia описал А.В. Горнов (2005). Появление всходов этого растения связано с местами роющей деятельности кабанов, которые могут уничтожать и локусы с генеративными растениями. В итоге популяции D. longifoiia могут иметь укороченные или удлиненные циклы сменяемости поколений, а полное отсутствие в экосистеме кабанов может привести к полному выпадению этого вида.
Онтогенетический спектр нормальной популяции в дефинитивном состоянии получил название базового, или характерного (Зауголъиоъь, 1974, Шоринаидр., 1976),так как он сохраняет свои черты в широком диапазоне условий. Л.Б. Заугольнова (1976) допускала существование нескольких (четырех) типов таких спектров: «базовыми» оказываются популяции с левосторонним спектром онтогенетических состояний, когда в популяции преобладают молодые особи (ювенильные - виргинильные), с правосторонним - с преобладанием в популяции стареющих особей (g3 - s). а также с одновершинным спектром с пиком на генеративных особях. Базовый спектр при таком подходе сводится к характерному набору спектров для данной популяции в некоторых фитоценозах и в некоторой амплитуде экологической обстановки. Возможно, целесообразно для таких спектров было бы сохранить приоритетный термин ^модальные» (от слова «мода* - наиболее часто встречающийся случай).
Уточнить данное понятие пыталась В.А. Черемушкина(2008). По ее мнению, базовый спектр - это модальный спектр, который отражает реакцию вида на типичную среду обитания, то есть раскрывает конкретную фитоценотическую позицию популяций данного вида, а характерный - это теоретически установленный спектр, зависящий от продолжительности онтогенетических состояний. На основе такого подхода у Dracocepftatom nufans характерным спектром она считала левосторонний, а базовым — центрированный.
Очевидно, «базовый спектр* в исходном понимании этого термина, если он вообще существует для данного вида растения, может быть только одним. Это приводит к необходимости введения понятия об идеальном онтогенетическом спектре популяции — характерном, согласно терминологии В.А. Черемушкиной. Такой идеальный онтогенетический спектр для конкретного таксономического вида растения может реализоваться при произрастании популяции в оптимальных эколого-ценотических условиях, когда продолжительность каждого возрастного состояния целиком определяется генотипом растения и не ускоряется, и не замедляется в случае отклонения этих условий от оптимума. Идеальный онтогенетический спектр может быть составлен на основании знания сроков пребывания особи растения в каждом из возрастных состояний. В абстрактном упрощении, если особь последовательно проходит 10 онтогенетических этапов и время пребывания ее на каждом из этапов одинаково, то в такой популяции будет по 10% особей каждого из этих онтогенетических состояний. В случае, когда срок пребывания особи в каком-то онтогенетическом состоянии по биологическим причинам в два раза короче (быстро проскакивает, например, фаза ювенильных растений), то доля ее в популяции тоже снизится в два раза и составит 5%. На основе такого подхода можно разрабатывать идеальный онтогенетический спектр для каждого вида растения. Однако его построение требует глубоких знаний о темпах онтогенеза изучаемого вида растения при его произрастании в оптимальных условиях. Такие данные имеются далеко не по всем таксономическим видам растений.
Л.Н. Бондаревой (2001) в работе, предшествующей публикации В. А. Черемушкиной, была разработана модель идеального онтогенетического спектра для семи видов луговых кормовых злаков (рис. 6.9). При равной продолжительности периода нахождения растений на каждом из онтогенетических этапов доля растений каждого этапа была бы одинаковой и составляла 11,1% (рис. 6.9А), С учетом реального времени продолжительности каждого из онтогенетических этапов идеальный онтогенетический спектр оказывается совсем другим (рис. 6.9Б). Этот подход требует де тального изучения биологии видов растений и занимает много времени, но он позволяет выявить оптимизированный идеальный онтогенетический спектр для каждого таксономического вида растений • Такой спектр может выступать в качестве эталона в сравнительных популяционных исследованиях.
В методическом плане ряд аспектов изучения онтогенетических спектров разработан еще далеко не полно. Главная проблема - отнесение конкретной особи к тому или иному онтогенетическому состоянию - не получила однозначного решения. Поливь- риантность онтогенеза при широкой амплитуде варьирования особей одного и того же онтогенетического состояния по виталитету часі о порождает серьезные ошибки. Избежать их можно, только отказавшись от излишней и ненужной дробности деления
|
11.1 |
11,1 |
И .1 |
11,1 |
11.1 |
11.1 |
11.1 |
11.1 |
11.1 |
|
і |
fm |
¦' I у j |
0? |
Os |
ss |
s |
||
|
A |
22,3 |
|||||||
|
18,5 |
16,6 |
|||||||
|
11,1 |
О. |
11,1 |
||||||
|
3,6 |
5,6 |
5.6 |
У |
0i |
ss |
5.6 |
||
|
I |
j Іт і |
*3 |
||||||
|
1 1 р |
1 ' |
s |
є
Рис. 6.9, Равновероятностный и реальный модельные онтогенетические спектры для семи видов луговых кормовых злаков:
А - доля особей в популяции в случае равной продолжительности жизни особи каждого из возрастных состояний %, Б - доля особей в популяции с учетом их реальной продолжительности нахождения в данном возрастном состоянии, % (по Л.Н. Бондаревой, 2001)
онтогенеза и приняв самостоятельные критерии для оценки онтогенетического и виталитетного состояния растений. Другая проблема, связанная с сезонными сменами состояния растений, тоже нуждается в методически однозначном решении с четким определением фенологических сроков для выявления онтогенетического спектра популяции. Так, у однолетних растений особи разного онтогенетического состояния существуют в разные периоды вегетационного сезона, а у многолетнего растения сныти обыкновенной Aegopodium podagraria семена прорастают в середине — конце апреля, и к середине лета волна семенного возобновления полностью спадает, остаются только ювенильные особи. Вето время в популяции исчезают всходы. Как же выявлять такие онтогенетические спектры? Тем не менее в большинстве публикаций по онтогенетическим спектрам популяций отсутствуют указания на сроки сбора материала.
Онтогенетические спектры популяций растений имеют самостоятельное значение, они почти не коррелируют с плотностью популяции и численностью растений в ней. Хотя в отдельных случаях такая корреляция может существовать. Она, например, выявлена у майника двулистного Maianthemum bi folium (Сулейманова, 2006).
Индикаторами для оценки статуса фитопопуляций оказываются и их фенологические спектры, но в последние годы этому простому и эффективному методу анализа популяционной жизни растений уделяется, к сожалению, мало внимания, хотя он представляет собюой особую форму биоразнообразия - ритмологическое внутрипопуляционное биоразнообразие. Е.В. Акшен- цев (2008), изучив феноритмологические спектры в популяциях Trollius europaeus, обнаружил высокую фенологическую гетерогенность популяций этого растения, которую он определил как один из механизмов популяционной устойчивости.
В целом, исследование закономерностей сменяемости поколений в данной популяции на протяжении достаточно длительного времени гораздо информативнее для оценки статуса и перспектив развития данной популяции, чем разовая и, в сущности, чисто случайная регистрация онтогенетического состава популяции. К тому же сменяемость поколений и формирующаяся онтогенетическая структура популяции могут реализовываться при разном жизненном состоянии особей растений, которые составляют популяцию. Это привело к необходимости независимого определения жизненного состояния особей и жизненного состояния популяций.
Еще по теме Периодизация онтогенеза:
- ГЛАВА 7 ПЕРИОДИЗАЦИЯ ОНТОГЕНЕЗА
- 7.6. ЭМБРИОНАЛЬНОЕ РАЗВИТИЕ МЛЕКОПИТАЮЩИХ И ЧЕЛОВЕКА 7.6.1. Периодизация и раннее эмбриональноеразвитие
- Эволюция онтогенез
- 7.1. ЭТАПЫ. ПЕРИОДЫ И СТАДИИ ОНТОГЕНЕЗА
- §1. Понятие онтогенеза
- Эволюция онтогенеза
- Онтогенез — основа филогенеза
- Эволюция стадий онтогенеза
- Целостность и устойчивость онтогенеза
- Эпигенетическая регуляция онтогенеза
- 13.2.3. Онтогенез — основа филогенеза
- Эмбрионизация онтогенеза
- Генетическая регуляция онтогенеза
- Автономизация — главное направление эволюции онтогенеза