Экосистема и ее структура
Под экосистемой понимается совокупность взаимодействующих абиотических и биотических компонентов в какой-либо ограниченной области. Абиотическая матрица (местообитание) обозначается как биотоп, а живая составляющая — как биоценоз.
Эта сильно упрощенная схема разъединяет то, что само по себе неразделимо, поскольку биоценоз постоянно изменяет и создает биотоп; даже его первоначальное пребывание создает свойственное ему жизненное пространство (биотоп), поскольку в абсолютно неживой форме его не бывает. Еловые леса ведь не растут изначально на свойственных им лесных почвах, но такие почвы формируются там, где достаточно долго росли ели. Эта дихотомия — биотоп-биоценоз символизирует тем не менее изменения в физико-химических условиях вместе со связанными с ними организмами.- Структура биоценоза
- Иерархическая структура
Биоценоз — это совокупность всех живых организмов экосистемы: растений, животных, грибов и микроорганизмов. Для совокупности всех растений существует понятие фитоценоза, или растительного сообщества (англ. plant community). Каждый вид такого «товарищества» (фитоценология, см. гл. 14), как правило, представлен многими особями, совокупность которых именуется популяцией (размножающееся товарищество). Популяция включает все возрастные состояния вида, а также те особи, которые присутствуют незаметно в почве в виде семян. Популяция состоит из генетически различных особей или генетически различных групп индивидов, возникших клональным способом (генетически тождественных в пределах клона), — триб (англ. ramets). При этом индивидуум (особь) воплощает собой определенный генотип или в случае генетически идентичной отдельной раметы — часть генеты (англ. genet — синоним клона). Поскольку возникшие из семян особи также могут быть генетически идентичны (апомиксис, см.
10.1.3.3), клоны, содержащие генетически однородные особи, встречаются чаще, чем можно было бы предположить исходя из их морфологии (например, апомиктический вид Taraxacum officinale). Аналогичная иерархическая структура имеется и в сообщес твах животных, грибов и микроорганизмов.- Таксономическая структура
Характер экосистемы создает присутствие определенных видов растений. По ним можно судить о местных условиях существования, поэтому часто в начале анализа экосистемы устанавливается список видов (индикаторные виды, см. 12.5.2.2; индикаторное значение, см. 14.3.3). Выявление числа видов и относительного обилия (частоты) отдельных видов — важная биологическая инвентаризация, обозначающаяся в фитоценологии (см. 14.3.3) немецкоязычных стран как структура сообщества (Bestandesstruktur). Поскольку английское слово «structure» (структура) почти исключительно используется для обозначения пространственной структуры (см. 12.5.1.5), целесообразно применить здесь понятие «видовой состав». Понятие биоразнообразия обычно применяется для количественной оценки числа видов. Тем не менее оно также включает внутривидовое генетическое разнообразие и надвидовой уровень, а именно многообразие растительных сообществ (см. 14.2.4.1).
- Функциональная структура
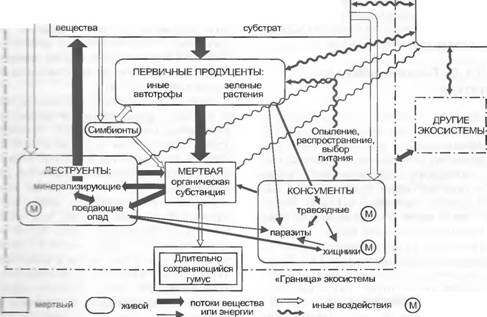
Все фотосинтетически активные организмы объединяются понятием первичных продуцентов. Им противостоят консументы (поедающие живые организмы) и дестру- енты (редуценты, разлагающие органическое вещество) Консументы функционируют непосредственно как травоядные животные (фитофаги, питающиеся растениями) или опосредованно как хищники (первого, второго или третьего порядка). Мертвые органические остатки в конце концов разлагаются самыми различными деструентами. К ним в особенности относятся детритофаги (детрит — мертвый опад; поедающие опад клещи и черви) и минерализаторы (бактерии и особые грибы). Такие пищевые цепи, или лучше пищевые сети (англ. food web), связывают друг с другом членов каждой экосистемы, как и потоки энергии и круговорот веществ.
Эти связи могут быть обратными и делают возможной в ограниченном объеме самореіу- ляцию, направленную против внешних изменений (рис. 12.4).Внутри растительного сообщества можно различать функциональные группы, или функциональные типы (англ. functional types). Теоретически их так же много, как и видов (см. введение к гл. 12), если же рассматривать функционально очень различающиеся возрастные состояния одного индивидуума (например, проросток — дерево), то их даже больше, чем видов. Концепция функциональных групп направлена на то, чтобы свести часто огромное разнообразие видов к небольшому числу «сходных по функционированию» категорий, что необходимо прежде всего для моделирования экосистем и теоретических обобщений (генерализации). Существует много таких попыток группирования; приведем некоторые важные. Наиболее простая и, пожалуй, наиболее используемая — это группирование по морфотипам (травы, кустарники, деревья или глубоко укореняющиеся в сравнении с поверхностно укореняющимися розет- кообразуюшие граминоиды и т.д.) и фенотипам (однолетние, многолетние, летнезеленые, вечнозеленые и т.д.; фенология. см. 7.7. и 12.3.1). При группировке по физиотипам исходят из особенностей обмена веществ, как, например,
ЧЕЛОВЕК как составная часть системы
12.4. Концепция экосистемы (по Н. Ellenberg).
схема совершенной экосистемы, т.е широкой саморегулирующейся структуры взаимо- зых организмов и окружающей среды (объяснения см. в тексте)
- "7 —6.5.9), по потребности в свете соответственно положению в сообществе (теневые и солнечные растения), специализации к субстрату (кальцефилы и кальиефобы) или свойствам резистентности (к засухе, засолению почвы, морозу. жаре). Симбиотипм основываются на способности к симбиозу с азотфиксиру- ю 'ими бактериями или специальными микоризными грибами. Критериями для объединения в группы могут служить и различные другие формы мутуализма и паразитизма.
Концепция, изначально инспирированная зоологией, — это группирование но господствующим жизненным стратегиям (англ. life strategies), ставшее классическим разделение растений по образу жизни на г-стратегов (первая буква слова «reproductive», что подчеркивает их устойчивость) и К-стра- тегов (первая буква слова «kompetitiver», что подчеркивает их преимущественно вегетативный способ размножения); с течением времени было развито много различных концепций стратегий растений. Особенно известна система треугольника Грайма, или CSR-стратегов (конкуренты, стресс-толе- ранты, рудералы). Она выводится из двумерной матрицы, в которой растения группируются по их толерантности к стрессу и нарушениям (малым или большим); одна из четырех комбинаций (сильный стресс и много нарушений) рассматривается как несуществующая, и остаются три категории. Они маркируют в углах треугольника экстремальные ситуации. Каждое растение занимает свое место между этими экстремами. Расстояния от угловых пунктов называются радиусами; например, стресс-радиус свидетельствует о степени стресс-толерантности вида. Как и каждая попытка типизировать многообразие жизненных ситуаций, эта концепция также критиковалась как слишком упрощенная.
П. Грабб (P. Grubb), напротив, считает, что категория стресс-толерантных растений должна быть отнесена к трем различным стратегиям, поскольку их поведение в течение жизни может изменяться. Ученый различает виды, которые сохраняют свой тип стратегии на протяжении жизни от всходов до взрослого растения (low flexibility strategy — стратегия низкой пластичности), виды, которые только в состоянии всходов хорошо переносят стресс (switching strategy— переключающаяся стратегия), и наконец виды, которые могут перестраиваться и в известной степени менять тип и переходить в другую категорию, когда условия очень благоприятны (gearing strategy — приспособительная стратегия).
Признаками стресс-толерантности наряду с другими являются медленный рост, многолетние органы, низкое репродуктивное усилие, сравнительно толстые листья с низким содержанием азота.Перечень примеров типизации и группирования можно продолжать еще долго. Опыт показал, что функциональные группы часто обнаруживают предполагающиеся для них общие функции только очень ограниченно и видоспецифические различия в пределах функциональных групп часто значительнее, чем между самими группами. При г руппировании может оказаться так, что особи разного возраста будут отнесены к различным функциональным группам, что затрудняет попытки группирования видов. Для разных целей предпочитают и разные способы группирования или даже создают новые группы (например, группы видов, устойчивых к озону, тяжелым металлам или застойному увлажнению). Морфо- и фенотипы могут по-прежнему представлять собой наиболее практичные функциональные группы для нетаксономического описания структуры фитоценоза.
- Материальная структура
Для наименования и разделения растительной субстанции в экосистеме установлены международные правила (все данные без исключения приводятся в сухом весе после высушивания при 80—100°С). Живая, надземная и подземная растительная составляющая экосистемы называется биомассой растений. В биомассу включаются также омертвевшие внутренние ткани (одревесневшие структуры), которые все же относятся к живому растению. Наружные отмершие части растений (надземные и подземные) обозначаются как некромас- са (англ. standing dead). Совокупность всех живых и отмерших наружных частей растений называется фитомассой. Ей противопоставляются отделившиеся мертвые части растений в экосистеме — подстилка (англ. litter), причем различают напочвенную подстилку (лежащую на поверхности почвы) и подземный опад (например, отмершие корни). Органические остатки, в которых простым глазом больше нельзя распознать структуру органа, относят к гумусовому комплексу (англ. soil organic matter, SOM), он охватывает все переходы от грубого гумуса до комплекса молекул гуминовых кислот в органической составляющей почвы.
Органическая субстанция животного происхождения (большей частью мелких почвенных животных) и микроорганизмов очень незначительна (lt; 0,1 %) и обычно прибавляется к углеродной составляющей почвы или вообще игнорируется, что, конечно, ничего не говорит о важной функции этих организмов в экосистеме.- Пространственная структура
Способ освоения воздушного пространства и почвы определяет не только общий внешний облик, но и свойства экосистемы. Морфология побегов и корней доминирующих видов растений, прежде всего геометрия или архитектура сообщества, придает каждой экосистеме свое неповторимое своеобразие, определяет, где происходит преобразование энергии, откуда поступают вода и питательные вещества.
Важнейшие признаки надземной структуры — высота доминантов, индекс листовой поверхности и вертикальное размещение листовой поверхности, подземной — типы корневых систем, максимальная глубина проникновения корней и вертикальное распределение корней в почвенном профиле.
Индекс листовой поверхности LAI (англ. leaf area index) — безразмерная единица, которая показывает, сколько квадратных метров листовой поверхности приходится на 1 м2 поверхности почвы (фактическая листовая поверхность независимо от ее ориентации в пространстве; при толстых или не плоских листьях это их наибольшая площадь проекции). В сомкнутых растительных сообществах па хорошо развитых почвах с достаточным водоснабжением LAI достигает значений 5 — 8. Для большинства посевных растений максимальное значение LA1 достигает почти 4. Плотность листовой поверхности LAD (англ. leaf area density), измеряемая в метрах в минус первой степени (м“‘), дается для всего сообщества в целом, если LAI разделить на высоту сообщества или на высоту отдельных ярусов (отношение величины листовой поверхности в м2 к 1 м3 объема сообщества). LAI и LAD определяют профиль освещенности в сообществе (см. 13.1.3).
Большая часть тонких корней находится обычно близко к поверхности почвы (lt; 1 м), часто даже в верхних 20 см почвенного профиля, там же сосредоточена большая часть минерализаторов и микоризных грибов. Часть корневых систем может достигать значительной глубины (см. табл. 13.3). Такие глубоко проникающие корневые системы служа! в основном для обеспечения водой. Ярусное распределение корней отдельных видов в почвенном профиле — классический пример дифференциации ниш, которая в значительной степени определяет биоразнообразие. Глубоко проникающие корни могут также служить для увлажнения верхних почвенных горизонтов (гидравлический лифт, см. 13.6.4). Многообразие типов корневых систем часто превосходит разнообразие типов побегов и является существенным структурным признаком каждой экосистемы.
Еще по теме Экосистема и ее структура:
- ГЛАВА 4. НАСЕКОМЫЕ В ЭКОСИСТЕМАХ
- 6. Биогеоценоз и экосистема
- 2. Экосистема села
- ВОДООБМЕН В БОЛОТНЫХ ЭКОСИСТЕМАХ
- БИОЦЕНОЗЫ.БИОГЕОЦЕНОЗЫ. ЭКОСИСТЕМЫ
- 6. АНТРОПОГЕННЫЕ ЭКОСИСТЕМЫ
- Аномалии экосистемы села
- Потребности и взаимодействия в экосистемах
- БИОГЕОХИМИЧЕСКИЙ КРУГОВОРОТ В БОЛОТНЫХ ЭКОСИСТЕМАХ[2]Н. П. Косых, Н. П. Миронычева-Токарева, Е. К. Вишнякова
- ВЗАИМОСВЯЗИ В ЭКОСИСТЕМАХ
- РАЗВИТИЕ ПРЕДСТАВЛЕНИЙ ОБ ЭКОСИСТЕМАХ
- Методологические подходы к изучениюструктурно-функциональной организации микробныхсообществ в наземных экосистемах
- ХИЩНЫЕ ПТИЦЫ И ИХ ЗНАЧЕНИЕ В ЭКОСИСТЕМАХ
- РОЛЬ ПОДЗЕМНЫХ ОРГАНОВ В КРУГОВОРОТЕ УГЛЕРОДАВ БОЛОТНЫХ ЭКОСИСТЕМАХ А. А. Титлянова
- ВЛИЯНИЕ ТРОФИЧЕСКОЙ И РОЮЩЕЙ ДЕЯТЕЛЬНОСТИ НА БЕРЕГОВЫЕ ЭКОСИСТЕМЫ
- КОСВЕННОЕ ВЛИЯНИЕ ЖИВОТНЫХ НА КРУГОВОРОТ ЭЛЕМЕНТОВ В НАЗЕМНЫХ ЭКОСИСТЕМАХ