Виталитет особей
Анализ виталитетной структуры популяции проводится в два этапа. На первом этапе устанавливается виталитет каждой из особей по репрезентативной выборке из популяции, на втором этапе на основании оценки доли особей разнрго виталитета вычисляется виталитетный спектр популяции.
Точность и объективность оценки состояния особей в витали- тетном анализе обеспечивается использованием количественных морфометрических параметров с вычислением их основных статистик: среднего арифметического, дисперсии, стандартной ошибки среднего арифметического и коэффициента вариации.
Концепция виталитета растений учитывает ряд важных общебиологических положений. Прежде всего, любой биологический организм является многопризнаковым объектом. Одна особь растения отличается от другой неисчислимо большим набором качественных и количественных особенностей — признаков.
В отличие от онтогенетических изменений, которые имеют однонаправленный характер, морфоструктурные особенности особей растений и их жизненное состояние обратимы: качество особей с течением времени в любом их возрастном состоянии может как улучшаться, так и ухудшаться, варьируя вокруг некоторого среднего значения, как видно из схемы на рис. 6.10, где показаны возможные направления и амплитуды изменений виталитета особей растений на каждом из этапов их онтогенеза.
При детальном изучении жизненного состояния растений ви- галитет особей следует определять для каждого онтогенетического состояния отдельно. При общих популяционных исследованиях обычно ограничиваются оценкой виталитета особей, находящихся в генеративном состоянии, когда их эколого-ценотическая роль максимальна.
Г
лг
¦УІ
-ы
/А
/ V
Яг
и-
\ А V
sm
Рис. 6.10. Варьирование виталитета особей растений на разных этапах онтогенеза:
- снижение виталитета; * + * - повышение виталитета.
Типичная амплитуда изменений уровня виталитетаПри разработке шкалы виталитета необходимо учитывать, что значение разных признаков, свойственных растениям, для раскрытия их жизненного состояния неодинаково. Использование одного или нескольких случайных признаков обычно не ведет к биологически значимым оценкам. Необходим отбор признаков с учетом их биологической и информационной ценности. В этой связи Ю.А. Злобиным (1980, 1989) был предложен алгоритм объективного выделения признаков, которые являются индикаторами жизненного состояния растений.
Набор признаков, который в наибольшей степени раскрывает жизненное состояние особей растений, получил название детерминирующего комплекса признаков. Для его выявления на объективной основе следует учитывать:
а) биологическую значимость каждого из признаков;
б) уровень варьирования признака;
в) скоррелированность признаков между собой;
г) вклад признаков в общую морфоструктуру растений.
Соответственно, для установления детерминирующего комплекса признаков был разработан алгоритм, включающий четыре этапа.
На первом этапе работы с растением новой жизненной формы из популяции делается репрезентативная выборка фитопопуляционных счетных единиц. Обычно ее объем составляет 35-50 и более растений. Для выявления темпов ростовых и формообразовательных процессов необходимы, как минимум, две выборки с интервалом в 10 дней. На втором этапе проводится полная морфо- метрия растений выборки с учетом не менее 20-30 параметров. Перечень таких параметров был дан в табл. 2.1. На третьем этапе проводится вычисление матрицы коэффициентов корреляции для полного набора признаков и на ее основе находятся корреляционные плеяды. На четвертом этапе проводится факторный анализ матрицы признаков растений.
При окончательном выборе признаков для анализа виталитета (обычно их выбирают три) учитываются структура корреляционных плеяд (выбираются признаки из разных плеяд), уровень изменчивости признака (предпочтение отдается признакам с наибольшим варьированием) и результаты факторного анализа (используют признаки, имеющие наибольший вклад в первый и второй факторы).
Рекомендуется использовать биологически и экологически важные и наиболее варьирующие признаки, наибольший вклад в факторные нагрузки по первому и второму факторам. Принятию правильного решения способствуют сводные схемы, подобные приведенной на рис. 6.11,Поскольку морфометрические признаки имеют разные амплитуды значений, А.Р. Ишбирдин и М.М. Ишмуратова (2004) предложили использовать их взвешенные значения. Для этого вычисляется индекс IVC:
IVC =— ,
N
где х - среднее значение признака в популяции;
х. - среднее значение признака для всех сравниваемых популяций;
N - число признаков.
Наибольшие значения IVC отвечают наилучшему состоянию. В этой связи отношение может являться оценкой размерной пластичности:
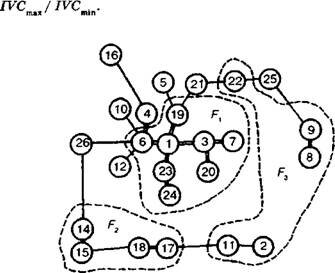
Рис. 611• Корреляционные плеяды и факторное решение для 26 морфометрических признаков чесночницы черешковой:
¦= - тесные корреляционные связи;
признаки, имеющие наибольший вклад в первые три фактора
В настоящее время детерминирующие комплексы признаков уже установлены для большого числа видов растений разных жизненных форм. Обычно первый признак - это фитомасса особи, второй - размер листовой поверхности, третий - репродуктивное усилие или семенная продуктивность особи.
Для оценки виталитета отдельных особей растений (фитопопуляционных счетных единиц) можно использовать одномерное, двумерное и многомерное (трехмерное) ранжирование. При одномерном ранжировании особи подразделяют на основе одного признака, двухмерном - двух, трехмерном - трех. Возможно и многомерное ранжирование по совокупности многих признаков. Ранжирование сводится к делению особей на три класса виталитета; высший (а), промежуточный (Ь) и низший (с).
Популяционное значение особей этих классов различно. Особи класса а обычно составляют основную группу растений популяции по фитомассе и обилию, класса Ь - группу размножения, а класса с - группу резерва, обеспечивающую контроль за размером экологической ниши, занимаемой популяцией. Особи классов а и б в наибольшей степени трансформируют среду обитания в пределах популяционного поля и препятствуют инвазии в него других видов растений. Особи класса с обычно бывают перегружены мутациями и обеспечивают соответствующие тренды микроэволюционного процесса. Так что в целом деление особей на классы виталитета по морфоструктурным признакам определяет не только их морфологические различия, но и популяционно-ценотические и экологические функции.В качестве алгоритма одномерного ранжирования (рис. 6.12) можно использовать следующие приемы;
а) весь набор значений, характеризующих величину того или иного морфопризнака - от наименьшего до наибольшего, разбивают на три равные части, находят граничные величины этих интервалов и, соответственно, подразделяют особи на три группы (процедураА);
б) с учетом соответствия набора значений признака нормальному статистическому распределению сначала выделяют группу промежуточных особей, соответствующих центральной зоне этого распределения (процедура Б): вычисляются значения t$x, и все величины признака, лежащие в интервале s + ts и s - ts‘, относят к промежуточному классу Ь. Эго осуществляется на ое-
|
класс с 0 ь с |
класс Ь g 1 § ! |
класс a —j p. |
||
|
^ lt; X ± fsx 0 1 |
I 5 1 |
gt;x±t$x |
||
|
lt; х ± ntsx |
gt; x і ntsx |
|||
Рис.
6.12. Способы деления общего интервала варьирования значений признака виталитета на частиковании статистики х ±t0M % , где х — среднее арифметическое, t - значение критерия Стьюдента при доверительном уровне в 95% ш,- ошибка среднего арифметического. В этом случае особи со значением избранного признака больше X +1;.Л5 составят высший класс а, а особи со значением признака менее х - ta05 % - низший класс с. Этой процедурой автоматически выделяется класс Ь. Но при низкой величине дисперсии признака класс b оказывается очень узким, и в таких случаях приходится прибегать к приему в);
в) ранжирование проводится в соответствии с процедурой В: в случае малых значений дисперсии признака величину tsj умножают на п, равное 2 или 3, а затем определяют ширину для промежуточного класса, как в способе Б, но применяется статистика x±nt0M % при п равном 2 или 3, что расширяет интервал промежуточного класса.
При двух- и трехмерном ранжировании (рис. 6.13) описанная процедура применяется для осей хну или х, у и z соответственно.
Следует особо подчеркнуть, что в качестве средней арифметической и дисперсии используются их значения, найденные для всех популяции и всех репрезентативных выборок из них, с которыми работает исследователь. Это позволяет как бы нанизывать на общую ось средних арифметических каждое растение и определять его статус.
|
II с |
1 с |
1 а |
|
Шс |
II Ь |
І Ь |
|
ШЬ |
III а |
II а |

Рис. 6.13. Графическая основа для группировки особей растений по трем классам виталитета: а - при двухмерном подходе; б - при трехмерном подходе
Одномерное ранжирование пока что наиболее часто используется в популяционных исследованиях. Применяется и двухмерное ранжирование, но наиболее информативным трехмерное ранжирование, оно позволяет одновременно учитывать признаки продукционного процесса, морфоструктуры особей и их участия в репродукции, то есть три важнейшие функциональные сферы растительных организмов.
Растения - многопризнаковые биологические системы, и редукция признаков, необходимая для любого сравнительного анализа, является очень сложной задачей. Помимо формализованного алгоритма, который был рассмотрен выше, всегда необходимо отдавать предпочтение таким признакам, которые наиболее важны для функционирования особей растений и обеспечивают устойчивое существование популяций.
Как правило, в популяционных и геоботанических исследованиях особи подразделяют на три класса жизненности, что вполне достаточно для практических целей. Принят такой подход и в виталитетном анализе.
Виталитетная структура популяций раскрывает соотношение в популяциях особей разного виталитета. В зависимости от преобладания в популяции особей разных категорий виталитета (а - высшего, Ь — промежуточного и с - низшего) определяется ви- талитетный тип популяции. Б.М. Миркин так оценил виталитет- ный анализ, сравнивая его с анализом онтогенетических спектров; «виталитетный подход... реализуется проще и часто дает не меньше информации, чем онтогенетический подход. При этом он более прагматичен, так как исследователь избавляется от необходимости субъективных домыслов о возрасте растений по их внешнему облику* (Миркин, Наумова, 1998: 95).
При проведении виталитетного анализа популяций долю особей того или иного виталитетного состояния принято выражать в долях единицы и представлять в виде гистограмм (Злобин, 1989). Всего теоретически существуют девять типов гистограмм, производных от равномерного статистического распределения (рис. 6.14). По соотношению в популяции особей разных классов виталитета популяции делятся на три базовые категории; процветающие, равновесные и депрессивные (табл. 6.5 и рис. 6.15). На основании равномерного статистического распределения, при котором в качестве нулевой гипотезы принимается равная вероятность частостей каждой из трех градаций виталитета особей, оказалось возможным предложить обобщенный индекс качества (виталитета) популяции (Q) в виде Q = 1/2(а + Ь), где а, Ь и с - доля особей высшего, промежуточного и низшего классов виталитета соответственно. Для процветающих популяций характерно соотношение - V2(а + Ь)gt;с, для равновесных - Уг * (а + Ь) * с, а для депрессивных - 7г • (а + Ь)lt;с. На рис. 6.14 этим трем категориям виталитета популяций растений соответствуют гистограммы:
Таблица 6.5. Классификация фитопопуляций по виталитетным спектрам
|
Витал итетное состояние особей |
Тип фитоценогюлуляции |
||
|
процветающие |
равновесные |
депрессивные |
|
|
Процветающие |
444 |
+ |
|
|
Промежуточн ые |
4 |
4 + + |
+ |
|
Угнетенные |
4 |
44-4 |
|

Рис. 6.14. Типы гистограмм
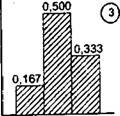
Рис. 6.14.2

Рис. 6.14.3

Рис. 6.14.4

Рис. 6.14.5
1-9 и соответствующие им кривые распределения частот, которые соответствуют основным категориям виталитетной структуры популяций. На оси абсцисс - градации жизненного состояния особей, на оси ординат - частости и относительные плотности. Над каждым столбцом гистограммы приведены значения теоретических частостей
Рис. 6.14.6
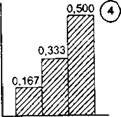
Рис. 6.14.7
процветающим - номера 2, 3 и 4, равновесным - 1,5 и 7 и депрессивным - 6, 8 и 9.
Статистическая достоверность отнесения популяции к той или иной их категории находится в соответствии с критерием х2. Для реализации всей совокупности этих расчетов имеется некоммерчес-
Рис. 6.15. Модельные спектры виталитетного состава популяций растений:
П - процветающие; Р- равновесные; Д- депрессивные популяции (а - особи наивысшего класса виталитета; Ь - особи промежуточного класса виталитета; с - особи низшего класса виталитета
кая компьютерная программа VITAL. Сопоставление спектров между собой удобно проводить по упомянутому выше методу JI.A. Жи- вотовского (1979). Виталитетный анализ целесообразнее всего проводить для группы генеративных растений, когда уровень жизнеспособности растений проявляется наиболее полно и контрастно.
Важная особенность виталитетного анализа состоит в том, что каждая популяция в итоге занимает некоторое «свое* место в ряду рассматриваемых популяций. Чем шире изучаемый эколого-фи- тоценотический градиент и чем больше охвачено популяций на нем, тем устойчивее будет оценка виталитета. Если число первоначально сопоставляемых популяций невелико, то добавление к нему очередной популяции будет вызывать смещение уже полученных оценок.
Виталитетный анализ с успехом используется в популяционных исследованиях разными авторами при работе с растениями разных жизненных форм. К настоящему времени виталитетная структура популяций изучена у более чем 100 видов растений: Dimeria neglecta (Семкин и др., 1995), Dactylorhiza romana (Кучер, 2000), Vaccinium myrtilius и Vaccinium uitis-idaea (Мазная, 2001, Коваленко, 2004), 13 видов луговых злаков и бобовых (Бондарева, 2005,Кирильчук, 2007), DiphasiastrumcomplanatumiYlamp;n- ченко, Чорноус, 2005), Cephalantkera rubra (Ишбирдин и др., 2005) и мн. др. (табл. 6.6).
Таблица 6.6. Основные.,убликациисрезультатамивиталитетногоана
лиэа популяций растений
|
№ П-'Р |
Объекты |
Авторы |
Дата публикации |
|
1 |
Подрост широколиственных древесных пород и неморапьные гравы |
Сухой И.Б. |
1986 |
|
2 |
Виды ее моральных лесных трав чз градиенте рекреации |
Баштовой Н,Г. |
1992 |
|
3 |
Calamagrostis langsdorffii |
Ким Г.Ю, |
1994 |
|
4 |
Origanum vulgare |
Троценко В.И. |
1994 |
|
С. |
Dimeria neglecta |
Семкин Б,И, и др. |
1995 |
|
6 |
Piantago major |
Козырева С. В. |
1998 |
|
7 |
Дер нови иные злаки |
Трубицына А.Н, |
1998 |
|
е |
Подрост широколиственных древесных пород |
Скляр В. Г |
1999 |
|
9 |
Amaranthus retroflexus |
Бариева Э.Р. |
1999 |
|
10 |
Dactylorhiza roman а |
Кучер Е. |
2000 |
|
11 |
Huperzia selago, Orthiilia secunda, Pyrota minor, Chimaphilla umbellate |
Панченко С.М. |
2000 |
|
12 |
Vaccinium myrtillus. V, vitis-idaea |
Мзэная Є.А. |
2001 |
|
ІЗ |
Восемь видев лесных трав |
Коваленко И.Н, |
2003 |
|
14 |
Восемь видов трав на урбанизированной территории |
Мельник Т.И, |
2005 |
|
15 |
Семь видов луговых злаков |
Бондарева Л.Н. |
2005 |
|
16 |
Травянистые многолетники Карпат |
Жиляев Г.Г. |
2005-2006 |
|
17 |
Diphasiastnjm complanatum |
Панченко С.М. др. |
2005 |
|
18 |
Cephalanther rubra |
Ишбирдин А.Р. |
2005 |
|
19 |
Rubus id3eus |
Быструшкина А. Г. |
2006 |
|
20 |
Cicborium intybus |
Олейникова Е М. |
2006 |
|
21 |
Valeriana tuberosa, V. dubia |
Ба рыщникова R И. |
2006 |
|
22 |
Шесть видов луговых бобовых трав |
Кирильчух Е.С. |
2007 |
|
23 |
Семь видов прикрепленных птолофитов |
Скляр Ю.Л. |
2007 |
|
24 |
CorydaBs bulbosa |
Фарадее ва М.Б. |
2008 |
По сравнению с анализом онтогенетического состава популяций анализ их виталитетной структуры имеет свои особенности и преимущества. Во-первых, виталитетный анализ выявляет первичные изменения в состоянии особей и популяций, которые предшествуют изменениям их онтогенетического состояния и тем более генотипа. Во-вторых, он наиболее чувствителен при выявлении особенностей эколого-цевотической обстановки, так как жизненное состояние при ее изменениях смещается в первую очередь. В-третьих, дает оценку особей и популяций в момент исследования, тогда как онтогенетические спектры отражают уже прошедшие воздействия на популяцию. И, в-четвертых, пригоден для анализа как одновозрастных, так и разновозрастных популяций.
Информационную ценность виталитетного анализа может иллюстрировать пример с анализом виталитетной структуры популяции клевера лугового Trifolium pratense при нарастании пастбищной нагрузки (рис. 6.16). Видно, что это ведет к снижению качества популяции клевера с падением индекса Q с 0,37 до 0,0 и снижением доли особей класса А с 60% на контрольном участке до их полного выпадения на участке луга с перевыпасом. Индикаторная роль виталитетной структуры популяций была показана при оценке уровня антропогенной трансформации природных экосистем Н.Г. Баштовым и Т.И. Мельник (1998) на техногенно- рекреационном градиенте для трех видов неморальных трав (Aegopodium podagraria, Mercurialis perennis, Polygonatum multiflorum). Выявлен переход популяций ряда видов лесных

Рис. 6.16. Виталитетная структура популяций клевера лугового за трех участках пойменного луга с разным режимом использования (по Е.С. Кирильчук, 2007)
кустарничков в депрессивную категорию в условиях промышленного загрязнения (Мазная, Лянгузова, 2008).
С виталитетной структурой популяций связано много других биологических феноменов. Г-Г. Жиляев (2004) на примере ряда видов травянистых растений продемонстрировал глубокие различия феноритмов в зависимости от жизненного состояния растений, которое оценивалось по трехбалльной шкале (рис. 6.17). При одинаковой продолжительности вегетации у белоуса торчащего Nardus stricta от Ж1 к ЖЗ плодоношение растений смещалось почти на месяц. Вообще, особи низкого жизненного состояния цветут позже у всех видов растений. Г.Г. Жиляев (2008) подчеркивает, что низкий виталитет не всегда является негативным показателем состояния растений и популяций. Так, у солъданеллы венгерской Soldanella hungarica и подбельннка альпийского Homogyne alpina в экотонных местообитаниях Карпат особи низкого виталитета переходят в состояние временного нецветения и поддерживают высокую плотность популяции, предотвращая вне-
VZZZ2 - выход в трубку ЯН - колошение
- бутонизация
- цветение
- осыпание плодов

Рис. 6.17. Сезонное развитие белоуса торчащего в зависимости от жизненного состояния особей (по Г.Г. Жиляеву, 2004)
дрение в популяционные поля других видов растений. Можно предположить, что во многих других случаях растения низкого виталитета играют важную роль в качестве резерва для восстановления популяции после воздействия различных стрессовых факторов.
Виталитетный анализ является полезным при рассмотрении механизма сукцессионных изменений фитоценозов. Это демонстрируют исследования Л.Н. Бондаревой (2005) семи видов злаков и Е.С. Кирильчук (2007 ) по отношению к шести видов бобовых луговых трав на пастбищном и сенокосном градиентах. Н.И. Став- рова и др. (2008) установили закономерности изменения виталитетной структуры популяций ели сибирской Picea obovata и березы пушистой Betula pubescens в ряду сообществ послепожарной сукцессии.
Достаточно сложным и специфическим может оказываться соотношение виталитетной структуры популяций и популяционной плотностью. У дрока красильного (Genista tinctoria) и ракитника русского (Ckamaecytisus ruthenicus) виталитет популяций возрастал по мере увеличение плотности, поскольку в этом случае обеспечивалось лучшее развитие симбиотических организмов на корнях этих бобовых растений (Гаврилова, Жукова, 2008).
Информационную ценность имеет виталитетный анализ при изучении агрофитоценозов как для культивируемых, так и для сорных растений, поскольку процветающие популяции обладают наибольшей фитомассой и выраженным эдификаторным действием.
Критерии отнесения популяций к тому или иному вяталитет- ному типу требуют дальнейшего уточнения, так как работ по выявлению соответствия устойчивого существования популяций в сообществах в течение многих лет с виталитетным спектром пока еще очень мало. Установлено только, что высокое виталитетное разнообразие особей в популяции не следует трактовать как признак ее устойчивости и процветания. Таким признаком является доля процветающих особей в популяции. Как правило, витали- гет популяций доминирующих видов всегда выше, чем виталитет ассектаторов.
При использовании виталитетного анализа в любой его модификации следует иметь в виду, что по своему содержанию оценки виталитета являются сравнительными и сохраняют свое значение только в пределах той группы популяций, для которой они проводятся. Пополнение группы новыми популяциями уточняет оценки виталитета, как было подчеркнуто выше. В идеале стабильные оценки виталитета могут быть получены ори включении в ряд сравниваемых популяций популяции, находящейся в условиях полного эколого-фитоценотического оптимума, когда генетический потенциал ее особей по росту, формообразованию и репродукции раскрывается полностью, и популяции из наиболее жестких условий произрастания, находящейся на пределе полного вымирания.
Виталитетный спектр позволяет прогнозировать динамические смены в популяциях растений. Например, регистрация процветающих популяций инвазионного сорняка, такого, как амброзия, указывает на необходимость разработки мер по предотвращению дальнейшей инвазии этого вида, а установление депрессивных популяций для редких охраняемых видов растений требует пересмотра мер охраны этих растений со сменой на более эффективные.
В целом, развитие концепции виталитета растений привело к пониманию необходимости различать как самостоятельные явления жизнеспособность видов, популяций и жизненное состояние особей. Виталитетный анализ - мощный инструмент популяционного анализа, который позволяет оценить как жизненное состояние особей популяции, так и виталитет популяции.
6.6.4. Размерная структура
Частным случаем виталитетной структуры популяций является размерная структура. Размерная структура характеризует соотношение в популяции особей разного размера. Размерная иерархия в популяциях возникает в процессе роста и развития растений и, как правило, бывает хорошо выраженной. Особи растений могут варьировать по размеру в силу разных причин. Анализ размерной структуры популяций растений ориентирован прежде всего на эколого-ценотический статус популяций, поскольку размер особей в первую очередь характеризует объем пространства, который они занимают и контролируют в экосистеме.
Размер особи оценивается обычно только по одному структурному признаку, который выбирается субъективно. Лучшей оценкой размера особи, безусловно, является величина ее общей или надземной фитомассы. При такой оценке размер оказывается важным биологическим признаком. Как было показано в ряде работ (Samson. Werk, 1986, Marba, 2007 и др.), в мире растений в целом от размера особи зависит продолжительность онтогенеза, конкурентная устойчивость особи, опыляемость цветков, величина репродуктивного выхода и много других важных свойств.
Часто исследователи характеризуют «размер растения» его высотой, диаметром розетки прикорневых листьев и др.
Критическим местом анализа размерной структуры популя ции остается выбор критерия оценки размера особей. В силу этого размерная структура популяции - понятие крайне неопределенное из-за субъективности выбора размерного признака и метода его учета.
По результатам учета принятого размерного признака особи ранжируют в восходящем порядке. В каждой популяции формируется континуум размера особей, который описывается либо нормальным статистическим распределением, либо распределением с выраженной правосторонней или левосторонней асимметрией, когда в популяции преобладают либо мелкие, либо крупные особи.
Обычно при анализе размерной структуры популяций особи делят на три или четыре размерных класса. Польские исследователи (Andrzejewska, Falinska, 1983) по соотношению размера особей в популяциях предлагали их деление натри группы:
- популяции, имеющие контрастный состав и состоящие только из крупных и мелких особей;
- популяции с асимметричным распределением особей при преобладании либо крупных, либо мелких особей:
- популяции с соотношением особей, соответствующим нормальному статистическому распределению, когда в популяции лучше всего представлены особи промежуточного размера.
Но эта система при ее разумности не нашла широкого применения в популяционной экологии.
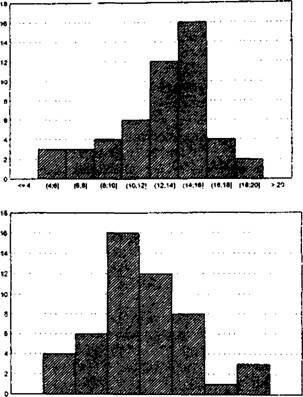
В ряде случаев, особенно при экспресс-авализе состояния популяций, изучение их размерной структуры оказывается полезным. Например (рис. 6.18), у подорожника большого Plant a go major анализ размерной структуры популяций всего по одному признаку - диаметру розетки - позволил установить своеобразие популяций в разных частях города. Н.И. Ставрова и др. (2000), изучая размерную структуру деревьев ели сибирской Picra obuaata н березы пушистой BetulaриЬе.чсепп, установили закономерно--1 и
о
Рисунок
Диаметр розетки, ом
Диаметр
¦ 1 (З.ч lt;*Ч («Л lt;*;*| «ОЩ lt;11,131 (14,13) Диаметр
І1.ІІ lt;Э;4) lt;6.е| (7.0) (0;iq (11;1Ц (13,14) gt;16 РОЗЄТКИ, СМ

Рис. 6.18. Размерная структура популяций подорожника большого в разных городских местообитаниях
изменения размерной структуры их популяций по мере формиро- вания древостоев: выровненность размера растений снижалась и возрастала асимметрия распределений.
По существу, анализ размерной структуры является прагматически редуцированным анализом виталитетной структуры популяций, которая более точно отражает дифференциацию особей по признаку жизненного состояния, чем размерная структура. В любом случае поликритериальный подход дает более объективную и ценную информацию о популяции, чем монокритериаль- ный, как это показал Е.В. Акшенцев (2005) при изучении популяций купальницы европейской Trollius europaeus.
- Основы методики изучения структуры популяций
При изучении структуры популяции первоначально закладываются и описываются стандартные геоботанические площадки. Их размер зависит от типа растительного сообщества и варьирует от 0,1-0,2 га для лесных сообществ до площадок в 25-10 мг для луговых сообществ. Сбор собственно популяционного материала проводят на площадках меньшего размера, которые закладывают в пределах основных площадок или рядом с ними. Размер таких площадок зависит от размеров растений и может составлять от 100 см2 при изучении популяций мелких мхов до 25 м2 при изучении крупных растений. Считается, что таких площадок должно закладываться 20-60 шт.
На популяционных пробных площадях учитывают целый ряд показателей, которые характеризуют популяцию: число особей, их размещение и др. Отбор особей для дальнейшего анализа проводят в случайном порядке в количестве 100-400 растений. У отобранных растений в зависимости от целей исследования определяют календарный возраст, онтогенетическое состояние и затем в лабораторных условиях проводят их полный морфометрический анализ. Набор необходимого оборудования небольшой. Это линейка, штангенциркуль, точные весы. При переводе весовых показателей в сухой вес растения необходим сушильный шкаф. Естественно, что виталитетный анализ выполняется для групп особей, которые находятся в одинаковом онтогенетическом состоянии.
lt;-4 (4Я1 (8:8) ftiei (10.121 (12:14) (14:18) (18:18) (1«0) gt;20


Диаметр
(•Л) (е;«Ч (10:12) (12:14) (14;16) (18,18) * 18 рОЭвТКИ, СМ
"1 014 . (4;Ч lt;*;7) (ЮН) (12:13) (14:18)
(1;2| (3:41 (8:8) (7:8) lt;9:10) (11:12) (1114) gt; 16 рОЭвТКИ, CM

Рис. 6.18. Размерная структура популяций подорожника большого в разных городских местообитаниях
изменения размерной структуры их популяций по мере формирования древостоев: выровненность размера растений снижалась и возрастала асимметрия распределений.
По существу, анализ размерной структуры является прагматически редуцированным анализом виталитетной структуры популяций, которая более точно отражает дифференциацию особей по признаку жизненного состояния, чем размерная структура. В любом случае поликритериальный подход дает более объективную и ценную информацию о популяции, чем монокритериаль- ный, как это показал Е.В. Акшенцев (2005) при изучении популяций купальницы европейской Trollius euro рае us.
Еще по теме Виталитет особей:
- Численность особей в популяции
- Целостность особей растений
- Вариабельность рабочих особей
- Биоразнообразие особей растений в популяциях
- Предпосылки возникновения внутрипопуляиионного разнообразия особей
- Горизонтальная структура - типы распределения и размещения особей
- ГЛАВА 7. РОСТ И РАЗВИТИЕ ОСОБЕЙ ПЧЕЛИНОЙ СЕМЬИ
- 7.4. Способность животных к оценке знаний и намерений других особей {^theory of mind»)
- Виталитетная структура
- Вертикальная структура
- Фитоиенотический оптимум популяиий и комплексная оиенка их устойчивости
- Демографические таблицы
- Что такое фитолопуляиия
- Индивидуальный и групповой отбор
- Популяция
- Математическое моделирование динамики популяций
- Родригесская летучая лисица Pteropus rodricensis Dob son, 1878 (V, 53)