Фотосинтез - световое воздушное автотрофное питание растений
Главная физиологическая функция листьев состоит в осуществлении фотосинтетического процесса. Образующиеся органические вещества являются основой для построения растения, у культурных растений часть его отчуждается человеком в форме урожая Происходит такое отчуждение и в естественных сообществах, где оно осуществляется фитофагами, начиная от мелких насекомых к заканчивая крупными травоядными животными.
Органические вещества при фотосинтезе образуются заново из неорганических веществ по схеме:
600, + 12Н,0 + 962 ккал - С^Н^О, Г 60, + 6Н,0,
Таким образом, фотосинтез относится к автотрофному типу питания живых организмов.
В процессе фотосинтеза последовательно решаются две самостоятельные биохимические задачи:
- поглощение солнечной радиации и использование ее энергии для расщепления молекул воды с получением во до рода-восстановителя;
- усвоение из воздуха углекислого газа, его восстановление еlt; лоро дом и последующий синтез первичных органических Веществ кз восстановленного углерода.
Получение водорода-восстановителя у всех зеленых растений осуществляется одним и тем же способом. Механизм ЭТОГО про цесса был изучен Хиллом и получил название реакции Хилла.
В противоположность этому усвоение углекислоте ’ аза 6а руется на разных биохимических механизмах, и в соответстви :gt; сущностью этих механизмов и строением их листьев зеленые растения подразделяют на две категории: С,- и С4-растения.
Реакции Хилла. Реакции Хилла осуществляются в гранулах хлоропластом и протека ют только на свету. Их упрошенная приведен: участие одновременно две пигментныефотосинтетнческне системы - первая (ФС1) И вторая (ФС2). Каждая из фотосистем гіоі тощает по одному кванту солнечного сьета, которые «выбивают» по одному электрону из молекул хлорофилла, передавая им свою энергию И переводя электроны ка возбужденный ьнергетичест 1-1
реализации реакций Ч'илле -ц* иь Мидекулы хлорофилла, благодаря потере электрон- ..
солнечного света
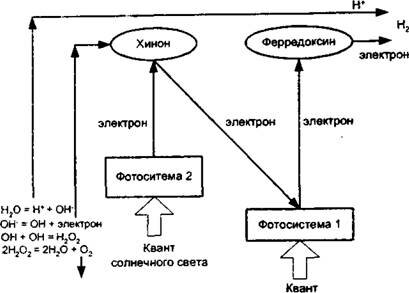
Рис. 3.2. Упрощенная схема световых реакций фотосинтеза
активизируются. Для замещения вакансии электрона молекула хлорофилла фотосистемы ФС1 просто забирает электрон от фотосистемы ФС2 и тем самым приходит в прежнее состояния, когда она вновь готова поглощать новый квант света.
Фотосистема ФС2 для замещения вакансия электрона осуществляет уникальный процесс: при обычной температуре в присутствии ферментов она расщепляет молекулу воды по схеме:
НгО = [Н+]+[ОН-].
Ион [ОН"] не очень прочно удерживает свой электрон, и фотосистема ФС2 отнимает его для замещения собственной электронной вакансии, переходя в прежнее состояния и восстанавливая свою способность к фотохимической работе.
Накапливающиеся радикалы [ОН], лишенные электрона, взаимодействуют между собой с образованием перекиси водорода:
[ОН) + [ОН]-Н202.
Особый фермент катадаза немедленно расщепляет ядовитую для клеток перекись на воду и кислород. Вода идет в биохимический процесс, а свободный молекулярный кислород 02 выделяется в атмосферу как побочный продукт фотосинтеза.
Оставшиеся ионы [ Н+] получают по электрону от промежуточного акцептора ферредокеина и превращаются в водород-вое становктелъ [Н2] - основной полезный продукт реакций Хилла.
Для осуществления реакций Хилла необходим ряд важных условий. Естественно, что они могут протекать при достаточном притоке солнечной энергии, то есть при оптимальной освещенью сти растений. Доступность воды реакции Хилла, как правило, не лимитирует, так как она расходуется в небольшом количестве. Зато для реакций Хилла необходима хорошая обеспеченность рядом минеральных элементов. В первую очередь это фосфор, который в форме АТФ участвует в переносах энергии, имеющих место пси фотосинтезе.
Магний входит в состав молекул хлорофилла, и позі ому хорошая обеспеченность им растений также необходима для оптимального фотосинтеза. Нужны и некоторые микрозле менты - железо, марганец, медь, которые входят в молекулы ферментов, обеспечивающих реакции Хилла. Поэтому обеспече ние растений популяции минеральными веществами является важным условием продукционного процесса.Сррастення. Сурастения распространены очень широко. К ним относится большинство выращиваемых сельскохозяйствен ных растений: пшеница, рожь, ячмень, картофель, сахарная свекла и мвогие дикорастущие. Блок-схема фотосинтеза Сурастений представлена на ряс- 3.3. В гранслах хлоропластов идут реакции Хилла, а в етроме - реакции цикла Кальвина.
й качеетзе поглотителя углекислого газа а цик ле ТСа.тьы- рыетупэе-т особый плтиатомный сахар в фоефоридироЕангк'й форме - рибулезадифосфат(РДФ)(рис. 3.4). Поглощая одну молекулу С02, каждая молекула РДФ преобразуется и превращается з две молекулы фосфогяицерлновой кислоты (ФГК). V." лег.тлы ФГК, в свою очередь, восстанавливают ся :я ¦: чет па ютпего из гранул водорода' восстановителя и превращают ли фосфсглицериновый альдегид (ФГА). Молекулы ФГА ислоль- зчются в пикте Кальвина по-разному. Одна их часть направляется на воссоздание РДФ. необходимого для поглощения новых погний ' т .7 е к и с л о г і г л з л. а другая иле г на синтез фр\ у: о л.-.. Ф р. кто за - это первый продукт фотосинтезе, и не ее ОСНОВ»- с связывания С02 базируется на фосфоэцол пировиноград ной кислоте (ФЭГГК), которая в присутствии фермента фосфоэнолпиру- ваткарбоксилазы очень активно «вылавливает* молекулы 002 из воздуха и реагирует с ними. Возникает четірехуглеродное соединение - щавелевая кислота. Щавелевая кислота благодаря работе специальных ферментов превращается в яблочную кислоту или аспарагиновую кислоту (если процесс идет в присутствии восстановленного азота).
Яблочная и аспарагиновая кислоты формируют своего рода депо связанного углерода и удерживают его в составе своих молекул.
Когда в цикле Кальвина возникает дефицит углерода для построения органических веществ, яблочная кислота расщепляется на С02 и пировиноградную кислоту. Углекислый газ передается в цикл Кальвина и там используется обычным путем, а пиро- виноградная кислота идет на воссоздание ФЭПК, которая вновь поглощает С02, и т.д. по циклической схеме.Таким образом, у С4-растений поглощение углекислого газа из воздуха осуществляют два акцептора; РДФ (цикл Кальвина) и ФЭПК (цикл Хетча - Слэка). Цикл Кальвина и цикл Хетча - Слэка пространственно четко разобщены. Цикл Хетча - Слэка протекает в обкладочных клетках проводящих пучков листьев, а цикл Кальвина - в обычных мелких клетках мезофилла листьев. Но биохимически эти циклы тесно связаны.
Биохимические особенности С4-растений дают им немало экологических преимуществ. Во-первых, ФЭПК гораздо полнее «вычерпывает* углекислый газ из воздуха, чем РДФ: РДФ прекращает связывание из воздуха углекислого газа при его концентрации 30-70 мкл/л, а ФЭПК продолжает поглощение до концентрации 10 мкл/л. Во-вторых, они устойчивее сохраняют высокий уровень фотосинтеза в стрессовых условиях, особенно в жару и во время засух. Фотосинтез у них может продолжаться и при закрытых устьицах за счет резерва углекислого газа, накопленного в форме яблочной кислоты. Все это в совокупности делает их более урожайными и продуктивными. Синтез органического вещества у С4-растений за сутки в среднем в 2-3 раза выше, чем у С - растений.
Еще по теме Фотосинтез - световое воздушное автотрофное питание растений:
- ВОЗДУШНОЕ ПИТАНИЕ РАСТЕНИЙ (ФОТОСИНТЕЗ)
- СВЯЗЬ МЕЖДУ УГЛЕРОДНЫМ (ВОЗДУШНЫМ) И МИНЕРАЛЬНЫМ (КОРНЕВЫМ) ПИТАНИЕМ РАСТЕНИЙ
- ПРИСПОСОБЛЕНИЯ РАСТЕНИЙ К СВЕТОВОМУ РЕЖИМУ
- ТИПЫ ПИТАНИЯ РАСТЕНИЙ
- Симптомы ЗАБОЛЕВАНИЙ, СВЯЗАННЫХ С НАРУШЕНИЕМ ПИТАНИЯ И СОДЕРЖАНИЯ РАСТЕНИЙ Симптомы нарушения произвольного питания
- МИНЕРАЛЬНОЕ (КОРНЕВОЕ) ПИТАНИЕ РАСТЕНИЙ
- Глава 4 ПИТАНИЕ РАСТЕНИЙ
- РАСТЕНИЯ И СОДЕРЖАНИЕ В ПОЧВЕ ВАЖНЕЙШИХ ЭЛЕМЕНТОВ ПИТАНИЯ
- ПИТАНИЕ РАСТЕНИЙ И ПРИМЕНЕНИЕ УДОБРЕНИЙ [42]
- Питание измененными частями растений.
- ИССЛЕДОВАНИЕ АЗОТНОГО ПИТАНИЯ И ОБМЕНА У РАСТЕНИЙ С ПРИМЕНЕНИЕМ ИЗОТОПА N15 [29]
- ВЛИЯНИЕ ВНЕШНЕЙ СРЕДЫ НА УСЛОВИЯ ПИТАНИЯ РАСТЕНИЙ И ЭФФЕКТИВНОСТЬ УДОБРЕНИЙ
- РОЛЬ РУССКИХ УЧЕНЫХ В РАЗВИТИИ УЧЕНИЯ О ПИТАНИИ РАСТЕНИЙ И НАУКИ АГРОХИМИИ
- ЗНАЧЕНИЕ ОТДЕЛЬНЫХ ХИМИЧЕСКИХ ЭЛЕМЕНТОВ В ПИТАНИИ РАСТЕНИИ