Взаимодействие между самками и родственные отношения между ними
Her недостатка в данных о том, что у моногинных видов самки настроены враждебно друг к другу и что рабочие особи наследуют эту враждебность, проявляя ее ко всем особям, за исключением родных и сводных сестер.
Благодаря этому семьи разделены значительными промежутками.- Взаимоотношения между самками
Враждебность между самками может на время ослабевать; например, у Apis mellifera и Formica fusca при замене старой самки молодой обе они некоторое время сосуществуют. На стадии, предшествующей роению, у медоносной пчелы семья вновь временно становится полигинной, но рабочие особи держат самок вдали друг от друга, как это бывает и во время деления семей у Eciton. Во всех этих случаях рабочие действуют заодно со своими самками и в их интересах; недавно, однако, у рода Leptothorax был описан интересный случай терпимости рабочих особей и враждебности самок (Provost, 1979,
- . Если две семьи одного из моногинных видов этого рода содержат в лаборатории, так что между ними имеется доступная той и другой арена, то в 40% случаев одна из них начинает двигаться в сторону другой, с тем чтобы соединиться с ней; при этом сначала переносятся расплод и рабочие особи, а под конец самка. Пришлая самка сразу вступает в схватку с „хозяйкой", используя в качестве оружия не мандибулы, а жало. Враждебность на короткое время передается и рабочим особям, которые хватают друг друга мандибулами, но не делают попыток ужалить, и драки между ними не бывают смертельными. Затем рабочие в течение длительного времени удерживают своих самок (это напоминает „фиксацию" молодых самок рабочими особями Eciton). Обе они оказываются прижатыми к субстрату, а потом одну из них (это всегда бывает „пришлая" самка) начинают медленно отпускать, а „хозяйку" разрывают на части и выбрасывают. Создается впечатление, что активная группа завоевывает пассивную. Рабочие смешиваются и облизывают новую самку, но неизвестно, привыкают ли они к двум наборам запахов, которые они различают, никак не проявляя этого, или же эта новая самка выделяет феромон, перекрывающий все другие запахи. У
Camponotus lateralis такое слияние семей оказалось невозможным; если удаляли одну самку, то слияние могло произойти в половине случаев, если же удаляли обеих самок, то семьи всегда сливались. Эти эксперименты подчеркивают важную роль самки в создании специфичности семьи. При наличии самки семья моногинного вида начинает защищать свое гнездо и расплод, хотя в лабораторных условиях она может кормиться вместе с чужими особями на общей арене.
Рабочие особи Apis mellifera заставляют репродуктивных самок прекращать драки; эти их действия известны под названием „balling" (свивание в клубок), и они нередко приводят к уничтожению той матки, которую рабочие не хотят признавать. Таким образом, „запечатывание", „фиксация" или balling может быть замедленным способом нейтрализации и наконец уничтожения самок, нежелательных для рабочих особей. Мы уже упоминали о соревновании между маткой и молодой самкой у Trigona subnuda. Благодаря участию рабочих особей весь процесс протекает более активно, чем у Leptothorax. Подобным же образом элиминируется избыток репродуктивных особей у термитов. После удаления экспериментатором пары особей-основателей наступает фаза замещения, во время которой производится очень большое число замещающих особей. Между ними возникают схватки, подробно описанные Liischer (1974); тех термитов, которые при этом бывают ранены, рабочие особи приканчивают, так что в конечном счете остается одна пара, принимающая на себя репродуктивные функции.
У термитов описано также несколько примеров слияния семей в природе; при этом после объединения двух семей одну из репродуктивных пар уничтожают, так же как самку у Leptothorax. По крайней мере у одного термита, Nasutitermes comiger, хотя это в общем моногинный вид, наблюдается факультативная полигиния.
В его семье может быть от 2 до 22 первичных самок (и самцов), которые мирно живут в многокамерном репродуктивном комплексе и все откладывают яйца. Как и у муравьев, эти полигинные семьи образуются в результате объединения отводков по инициативе самок, однако возможно и кооперативное основание семей. Thorne (1982) обнаружил такие полигинные семьи в районах Панамы в восстанавливающемся вторичном лесу. Очень интересные взаимоотношения существуют между самками у полигинных видов. У Camponotus herculeanus каждая самка имеет в гнезде собственную территорию (Holldobler,- . То же самое наблюдается у Bombus atratus (Sakagami el al., 1967a). Там, где нет такой территориальности, следовало бы ожидать иерархической структуры, основанной на доминировании. Скорее всего, это выражается в не слишком явной форме, и на положение данной особи в иерархии указывает та зона гнезда, в которой она находится, очередность в получении пищи и взаимоотношения с расплодом, как, например, у полигинной осы Metapolybia. В семьях Leptothorax curvispinosus число самок может достигать трех; они, по-видимому, относятся друг к другу дружелюбно и вместе откладывают яйца, но в каждой из двух групп, изученных Wilson (1974а, Ь), одна из самок относилась к собственным яйцам гораздо более заботливо, чем к яйцам двух других самок! Эта верховная самка „как бы случайно" раздавливала и поедала яйца других самок, а также рабочих особей, и можно было видеть стычки между ними из-за этих яиц, хотя рабочие, по-видимому, сами никогда их не едят.
Большую часть пищи самок составляет содержимое зоба рабочих особей, которое те им отрыгивают; кроме того, самки дополняют свой рацион веществами, которые они берут изо рта личинок. Рабочие, не имевшие корма, который они могли бы предложить самкам, избегали их или даже спасались от них бегством; они, по-видимому, узнавали самок, ощупывая их головы своими усиками. Быть может, эти самки занимали слишком низкое место в ряду по степени предпочтения со стороны рабочих особей.
Wilson не мог найти никаких указаний на то, что самки узнавали друг друга, хотя они явно отличали чужие яйца от своих собственных. Те случаи, когда верховной самке удавалось уничтожить яйца других самок, можно рассматривать как случаи функциональной моногинии,как, например, у Myrmecina graminicola (Baroni Urbani, 1968). Сходные ситуации были обнаружены у рода Formicoxenus и у вида Leptothorax gredleri (Biischirtger,- . У Lasius sakagamii-nonmmmoro вида, близкого к L. niger,- самки скапливаются в густо населенных зонах гнезда, отведенных для выращивания расплода, но в каждой такой зоне они обычно занимают разные камеры; только в период, когда происходит набор новых дочерних самок, в каждой камере может находиться больше чем по одной самке (Yamauchi et al., 1981).
Рис. 14.4. Перемещения самок Myrmica rubra в узкостенных садках в лаборатории. Передвижения самок регистрировались в течение 30 мин. Обратите внимание на обособление всех самок в разных зонах и на локальность их перемещений. Самки избегали друг друга и иногда даже вступали в драки. (Evesham, 1982.)
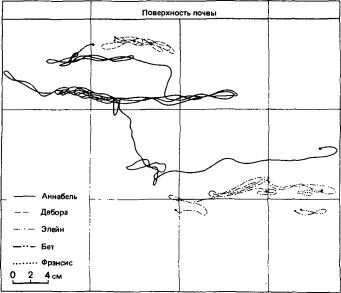
Изучение взаимоотношений между самками у Myrmica rubra показало, что хотя их в среднем бывает по 16 на семью (с диапазоном колебаний от 0 до более чем 70; Elmes, 1973), они не образуют скоплений, если не считать весны, когда они выходят наверх, чтобы согреться. У самок бывают „индивидуальные участки" (но не настоящие территории), в пределах которых они живут; самки сильно различаются по подвижности-одни ведут в основном сидячий образ жизни, находясь все время в камере в глубине гнезда, другие более активны, часто поднимаются наверх и даже выходят на поверхность почвы и убегают от гнезда (рис. 14.4). Встретившись друг с другом, такие самки вступают в короткую схватку, действуя передними ногами и усиками; изредка они пускают в ход челюсти, но никогда не используют жало. По-видимому, самки, ведущие сидячий образ жизни, больше привлекают к себе внимание рабочих особей и откладывают больше яиц; им оказывают предпочтение, их, вероятно, лучше кормят, и можно сказать, что они занимают в сообществе доминирующее положение, проявляя минимум агрессивности (Evesham, 1982).
У этого вида группы особей, лишь недавно оставшиеся без самки, охотно принимают самок из своей собственной семьи и даже из чужих семей. Степень предпочтения самки может быть различной: если предложить муравьям мертвую самку из другой семьи, то она будет отвергнута и вынесена в камеру, предназначенную для отбросов, тогда как самку из той же семьи, хотя и мертвую, муравьи вносят в гнездо и кладут на расплод, т. е. принимают. Живые чужие самки направляются в сторону камер с расплодом, а рабочие особи могут либо задержать их, либо позволить им остаться и откладывать яйца. Этих яиц, возможно, будет меньше, чем могла бы отложить самка, принадлежащая к их собственной группе, и их откладка не обязательно заставит рабочих начать откладывать кормовые яйца; фактически они в таких условиях откладывают значительную долю репродуктивных яиц (Brian et al., 1981b; Winterbottom, 1981). Таким образом, к этим самкам проявляется терпимость, но они не могут влиять на поведение рабочих особей и косвенно воздействовать на функцию их яичников. Группы рабочих, не имеющие самки в течение многих недель, не соглашаются принять даже самку, принадлежащую к их собственной семье, и относятся к ней, по крайней мере в первое время, слегка враждебно, держа ее в подчинении несколько часов или даже дней. У некоторых муравьев такая враждебность по отношению к самкам может даже выйти за пределы гнезда, распространяясь на окружающую территорию через посредство рабочих особей; это наблюдается, например, у Lasius niger и L.flavus, которые оба относятся весьма агрессивно к самкам собственного вида (Pontin, 1961b). Муравьи Pogonomyrmex, обнаружив на своей территории самок, уносят их на край гнезда, не причиняя им вреда, что еще чрезвычайно любезно с их стороны (Holldobler, 1976а)!- Степень родства между самками
Кажется вероятным, что высокая степень родства между самками (если, например, они сестры) способствует их признанию и дружественным взаимоотношениям в гнезде.
У Tetramorium caespitum самки сотрудничают между собой только в том случае, если они действительно приходятся друг другу сестрами (Poldi,- . В ряде случаев меченых крылатых самок обнаруживали впоследствии в их родительском гнезде и даже в близлежащих гнездах; это наблюдалось, например, у Myrmica sabuleti (Brian, 1972; Jones, личное сообщение) и М. rubra (Elmes, 1973). West-Eberhard (1969) и Klahn (1979) показали, что у ос живущие вместе самки-сестры, a Ross, Gamboa
- ,-что самки ос предпочитают объединяться с товарками по родному гнезду и способны к этому даже после пребывания в изоляции в течение 100 дней.
Metcalf и Whitt (1977а, Ь) на основании анализа изозимов установили существование сходной степени родства между самками-основательницами у Polistes metricus и обнаружили также много других интересных черт их общественной жизни. Семьи основывают одна особь или две сестры (или двоюродные сестры), каждая из которых может копулировать дважды и получить при этом смесь, в которой отношение спермы от двух самцов будет составлять 9:1. Доминирующая самка производит 78% всех имеющихся в семье самок и 87% самцов, а остальных самок и самцов производит подчиненная самка. Рабочие особи редко откладывают яйца, если самка жива; если же она погибнет, то одна из рабочих особей занимает доминирующее положение и откладывает 95% всех яиц. Инбридинг так редок, что им можно пренебречь, и коэффициент родства (г) между потомками репродуктивной самки равен 0,66, если в семье только одна самка, и 0,49-если их две. Коэффициент родства между репродуктивными самками и их матерью равен 0,50, если это доминирующая самка, и 0,32 - 0,36, если это подчиненная самка; таковы же коэффициенты родства самцов.
Гнезда, закладываемые двумя самками-основательницами, лишь в 2,25 раза превосходили по успешности развития гнезда, заложенные одной основательницей, но приспособленность доминирующей самки, оцениваемая на уровне семьи, превосходит таковую одиночной основательницы. Даже вторая самка, хотя она присоединяется к доминирующей самке и занимает подчиненное положение, функционирует не менее эффективно, чем если бы она начинала устраивать гнездо в одиночку! Тем не менее гнезда, в которых самок две, имеют больше шансов на успех и обычно терпят неудачу только в случае гибели второй самки до того, как рабочие особи в состоянии заменить ее; самки гибнут от хищников или от несчастных случаев, находясь вне гнезда.
Metcalf и Whitt приходят к выводу, что эти данные согласуются с объяснением возникновения общественного образа жизни на основе теории отбора родичей, но причины объединения самок не ясны, поскольку самки почти так же хорошо справляются в одиночку.
Дополни гельную информацию дает изучение рабочих особей и самок в семьях, живущих в естественных условиях, методом анализа изозимов'. Pearson (1980) изучал две популяции Myrmica rubra в соседних долинах; в одной из них не было обнаружено родства между самками, а в другой самки находились в родстве в одном году и не были родственны между собой в следующем году-удивительный случай изменчивости. У полигонного вида Myrmecia pilosula самки родственны друг другу примерно на уровне двоюродных сестер, а родство между рабочими особями весьма отдаленное (Craig, Crozier, 1979). Частоты генотипов в пределах каждой касты распределяются в соответствии с законом Харди-Вейнберга, т. е. позволяют предполагать аутбридинг, не сопровождающийся отбором.
В исследовании Crozier (1977), проведенном на Aphaenogaster rudis, однажды осемененная самка и ее потомки-рабочие оказались родными сестрами, а самцы происходили от самки
1 При этом методе фактически выясняется не степень родства, а степень сходства геномов, определяемая путем анализа изозимного состава изучаемых локусов. Отсюда и необычные выражения вроде „самка и ее дочери являются сестрами". При гапло-диплоидном определении пола генетическое сходство между матерью и дочерьми теоретически должно быть 0,5, между сестрами-0,75 (так как одна хромосома, полученная от отца, у них всегда одинакова), а между двоюродными сестрами-0,375. Но фактическое сходство может быть и выше, если генотипы матерей и отцов сходны,-Прим. ред.
или (если самки не было) от рабочих особей. У одной из форм Iridomyrmex purpureus (Dolichoderinae), у Rhytidoponera chalybaea и еще у одного вида (два последних вида-моногинные) рабочие особи приходятся друг другу родными сестрами (Crozier, 1980); у полигонного вида Rh. impressa рабочие особи связаны менее близким родством.
При изучении у муравья-«рабовладельца» Formica sanguinea в Финляндии одного ферментного локуса, кодирующего малатдегидрогеназу, было обнаружено расщепление по четырем содержащимся в нем аллелям (Pamilo, Varvio-Aho, 1979). Моногиния в данном случае исключается, так как 75% самок гетерозиготны (против ожидаемых 35%) ив 10 из 92 гнезд обнаружены рабочие особи, относящиеся к трем или четырем различным генотипам. Цитированные авторы полагают, что в каждом гнезде яйца откладывают несколько самок, связанных между собой родством; возможно, это пары мать - дочь, причем большую часть яиц откладывает мать. Каждая самка копулирует один раз. Географическая изменчивость у этого вида незначительна, и только 2,4% всей изменчивости относится к межпопуляционной. Методы разделения белков позволят расширить подобные генетические исследования.
Они дадут возможность изучать родственные связи и механизмы, используемые в сообществах.
Таким образом, организация общественных насекомых в „семьях" выражается в весьма разнообразных формах-от строго семейной моногинной группы до гораздо более рецептивной полигонной группы с низким коэффициентом родства между особями. Группа первого типа-это прежде всего приспособление для расселения, и она регулирует плотность популяции при помощи разного рода механизмов, связанных с территориальностью, хотя такая организация, несомненно, сдерживает социальных паразитов и других врагов. При высоких плотностях популяции самки- или пары-основательницы могут объединяться и регулировать производство яиц путем установления доминирования. Там, где утверждаются такие полигинные группы с циклическим замещением самок, между самками сходного типа существует определенная система разделения, и это, вместе со способностью рабочих особей выбирать и регулировать плотность самок, приводит к созданию популяции, экономически соответствующей имеющимся ресурсам. Сведений о деталях используемого для этого механизма и о том, насколько эффективно он поддерживает наилучших производителей, пока еще мало, однако нет сомнений, что общественная организация насекомых зависит от условий среды. Накапливается все больше данных в пользу того, что разведывательная моногинная группа по мере ее закрепления может перейти в полигинное состояние, сопряженное с конкуренцией.
Еще по теме Взаимодействие между самками и родственные отношения между ними:
- ПОГОДА И КЛИМАТ: В ЧЕМ РАЗНИЦА МЕЖДУ НИМИ
- 3.6.5.2. Взаимодействия между генами в генотипе
- Взаимоотношения животных между собой
- Конкуренция между видами
- Между различными видами
- Границы между сообществами и ценотическими системами
- Формы взаимоотношений между популяциями разных видов растений
- МЕЖДУ ДВУХ СТУЛЬЕВ
- ЧТО ОБЩЕГО МЕЖДУ БЕЛКОЙ И КАБАНОМ?
- Глава И ЭКОЛОГИЧЕСКИЕ СВЯЗИ НАСЕКОМЫХ МЕЖДУ СОБОЙ И С ДРУГИМИ животными
- Географические связи между ареалами
- Взаимосвязь между гидрогеологическими/>условиями и заболачиванием
- Конкуренция между особями одного вида
- Прививки между культурными сортами картофеля
- Прпвивки между культурным и диким картофелем
- 3.4.3.3. Ген — функциональная единица наследственного материала. Взаимосвязь между геном и признаком