Основные черты организации хордовых
Эмбриональное развитие всех хордовых сходно. Его примером может быть эмбриогенез ланцетника. Оплодотворенное яйцо испытывает полное дробление, в результате которого образуется полая шаровидная бластула (рис.
6, А). Более крупные клетки ее вегетативного полюса впячиваются внутрь (инвагинация), образуя двуслойную гаструлу; ее наружный слой — эктодерма, или кожный листок, внутренний — энтодерма (кишечный листок); полость гаструлы (ее называют полостью первичной кишки) открывается наружу гастропором (бластопором). Гаструла вытягивается, гастропор уменьшается. Далее на спинной стороне гаструлы обособляется нервная пластинка, затем она опускается, а примыкающие участки эктодермы приподнимаются в виде складок и смыкаются над нервной пластинкой, которая оказывается лежащей под эктодермой (рис. 6, Б). Края нервной пластинки припод-
Deuterostomfa (Deutencephalia)
Рис. 5. Предполагаемые схемы эволюции хордовых животных. А — по Баррингтону; Б — принятая в учебнике
А — продольные разрезы. / — бластула, II—IV — гаструляция, V—VI — образование мезодермы, хорды и нервной системы: / — анимальиый и 2 — вегетативный полюса, 3 — гастральная полость, 4 — гастропор, 5 — нервный канал, 6 — пейро-кишечный канал, 7 — невропор, 8 — складка мезодермы. 9 — целомические мешки, 10 — хорда, // — будущий рот, 12 — будущий задний проход;
Б — поперечные разрезы: / — эктодерма, 2 — энтодерма, 3 — мезодерма, 4 — полость кишечника, 5 — иервиая пластинка, 6 — нервная трубка, 7 — невроцель, в — хорда, 9 — целом (вторичная полость тела);
В — поперечный разрез через личинку: / — хорда, 2 — целой, 3 — гонотом, 4 — кишка, 5 — миотоы, 6 — кожный листок, 7 — склеротом, 3 — нервная трубка
I — нервная трубка, 2 — хорда, 3 — пищеварительная трубка, 4 — ее жаберный отдел — глотка, 5 — печень (печеночный вырост), 6 — сердце с сосудами
нимаются и смыкаются, образуя нервную трубку с полостью — невро- цслем.
На этой стадии невроцель открывается нервно-кишечным каналом в полость кишечника, а спереди через невропор — во внешнюю среду; оба отверстия позднее закрываются.Одновременно идет дифференцировка энтодермы. На спинной стороне она утолщается, образует желоб, который, отшнуровавшись от кишки, превращается в плотный продольный стержень — хорду (рис. 6, Б), лежащую над кишечником, непосредственно под нервной трубкой.
По бокам формирующейся хорды образуются мешковйдные выпячивания, вскоре отшнуровывающиеся от первичной кишки и превращающиеся в парный ряд метамерно расположенных целомических мешков. Их стенки представляют собой мезодерму (третий зародышевый листок), а их полости — зачатки вторичной полости тела, илн целома. Разрастаясь, целомические мешки подразделяются на лежащий сбоку хорды и нервной трубки толстостенный спинной отдел — сомит — и находящийся у кишечника брюшной отдел — брюшную пластиику. Каждый отдел — сомит и боковая пластинка — получают свой участок вторичной полости тела. В дальнейшем из сомитов образуются миомеры илн мускульные сегменты и соединительная ткань. Из последней возникают: оболочка вокруг хорды, в которой у позвоночных формируется осевой скелет (позвоночник); оболочка нервной трубки; перегородки между миомерами (миосепты) и соединительнотканный слой кожи — кориум, или кутис. Полости брюшных пластинок сливаются в общую вторичную полость тела — целом. Стенки боковых пластинок дают начало мускульному и соединительнотканному слойм кишечной трубки, листкам брюшины н брыжэйке. Из эктодермы образуется поверхностный (Эпителиальный) слой кожи, а из энтодермы — слизистая оболочка (эпителий) пищеварительной трубки и ее железы. На переднем конце тела зародыша путем прорыва образуется ротовое отверстие, а на заднем, в области заросшего гастропора, — анальное отверстие.
Характер эмбрионального развития видоизменяется у части рыб, У рептилий и птиц: в связи с резким увеличением в яйце питательных веществ происходит не полное, а лишь частичное дробление яйцеклетки и типичная шарообразная бластула не образуется.
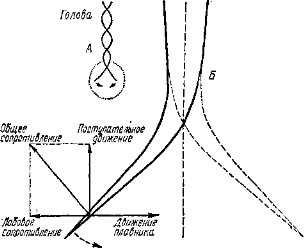
Двигательная система. Основой тела хордового животного служит особого тнла двигательная система — миохорд, или мышечно-хордаль- ный комплекс, образованный центральной опорной струной — хордой и прилегающей к ней метамер ной мускулатурой, состоящей из
мышечных сегментов — миомеров, разделенных соединительнотканными перегородками — миосептами. Тесная связь гибкого внутреннего скелёта с мускулатурой обеспечивает эффективное движение. Одновременно формируется веретенообразная форма тела, гидродинамически наиболее выгодная для активно двигающегося в воде животного (рис. 7).
Образование миохорда оказалось конструктивно удачным решением, предопределившим дальнейшую прогрессивную эволюцию хордовых. Его последующие перестройки (замещение хорды позвоночным столбом, усложнение мускулатуры и т. п.) при сохранении принципиальной схемы позволили повысить интенсивность движения и увеличили общую подвижность. Даже на суше осевой скелет с парными конечностями позволил эффективно преодолеть силы гравитации, обеспечив появления бегающих, прыгающих, роющих, лазящих и летающих позвоночных животных. Совершенствование двигательной и опорной функции миохорда способствовало заселению хордовыми животными (подтипом позвоночные) пригодных для жизни участков Земли: биотопов суши, включая почву, нижних слоев воздуха и практически всех водных биотопов. Одновременно шло увеличение размеров тела: по абсолютным размерам хордовые значительно крупнее животных других типов. Даже гигантские осьминоги и кальмары (из головоногих моллюсков) уступают по величине наиболее крупным рыбам, китам и ископаемым ящерам.
Биохимические особенности. Развитие двигательных способностей в типе хордовых животных обусловлено не только совершенствованием строения, но и изменением физиолого-биохимических механизмов. У всех животных необходимая для работы мышц энергия передается с помощью аденозинфосфорных кислот. Богатая энергией аденозинтри- фосфорная кислота (АТФ) отдает ее сократимым белкам мышц, превращаясь в обедненную энергией аденозиндифосфорную кислоту (АДФ).
АДФ вновь «заряжается» энергией, освобождающейся при постоянно идущих в организме процессах окисления (дыхания) и частично гликолиза. При продолжительной и напряженной работе мышц необходима постоянная «подзарядка» АДФ за счет веществ — фосфагенов, служащих как бы резервом (запасом) энергии (рис. 8).У большинства беспозвоночных животных таким веществом в мышцах служит аргининфосфат \ количество которого в организме сильно меняется, так как аминокислота аргинин используется при биосинтезе белков и у высокоактивных животных не может надежно обеспечить потребности организма. У хордовых животных, включая бесчерепных и большинство оболочников, фосфагеном служит креатинфосфат. Креатин образуется при распаде аминокислот (аргинина, метионина и др.) и выводится из организма с мочой, потом и слизью. Но, присоединяя богатые энергией молекулы фосфорной кислоты, креатин, аналогично аргинину, превращается в фосфаген, способный отдавать энергию для превращения АДФ в АТФ. Запасы креатина в организме
/|\
 alt="" />Н20 СОа NH3
alt="" />Н20 СОа NH3

NH2
Рис. 8. Схема биохимии мышечного сокращения у позвоночных животных (объяснения в тексте)
относительно постоянны и его использование в качестве фосфагена креатинфосфата представляет известное преимущество для животных с высокой подвижностью. Заметим, что у некоторых асцидий обнаружен лишь аргининфосфат. У полухордовых и некоторых морских ежей есть аргининфосфат и креатинфосфат, у офиур — только креатин- фосфат, у голотурий, морских звезд и большинства морских ежей — только аргининфосфат.
Встречается креатинфосфат и у некоторых, особенно свободноплавающих полихет. Использование в качестве фосфагена креатинфосфата в разных типах животного царства (хордовые, иглокожие, кольчатые черви) может служить примером конвергентной эволюции биохимических особенностей.Энергия жизнедеятельности и уровень подвижности определяются интенсивностью окислительных процессов, обеспечиваемых переносом кислорода дыхательными пигментами. У беспозвоночных последние содержат железо (эритрокруорины, гемэритрины, хлорокруорины) или медь (гемоцианины моллюсков, ракообразных и некоторых паукообразных) и обычно имеют высокую молекулярную массу: около 3 млн. эритрокруарин червя Arenicola, 6 млн. 680 тыс. — гемоцианин улитки Helix и т. д. У большинства низших хордовых (оболочники, бесчерепные), как и у многих беспозвоночных, дыхательных пигментов нет. Лишь у некоторых асцидий (оболочники) в крови находится зеленый пигмент гемованадин; содержание в нем ванадия может достигать 0,2% сухого веса асцидии и значительно превышает концентрацию этого элемента в морской воде. Функция гемованадина остается неизвестной; видимо, она не связана с переносом кислорода.
У всех позвоночных животных (за исключением нескольких видов антарктических рыб) дыхательные пигменты содержат железо: в эри-
троцитах крови — гемоглобнны, й в красных мышцах — миоглобины. Миоглобии представляет собой мономер с молекулярной массой 17— 18 тыс. Молекула гемоглобина построена из четырех таких мономеров (молекулярная масса 68— 72 тыс.). Миоглобин обеспечивает накопление запасов кислорода в мышцах и отдает его при недостатке 02 в мышечной ткани. Гемоглобин крови переносит запасы кислорода по всему, организму. В связи с этим кривые диссоциации (отдачи) кислорода у гемоглобина и миогло- бина отчетливо различны (рис. 9). Сочетание двух дыхательных пигментов обеспечивает устойчивое снабжение организма кислородом, особенно при значительных мышечных напряжениях.

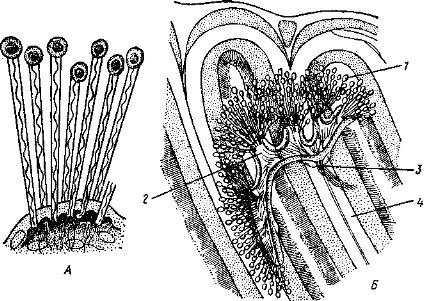
 Повышение уровня окислительных процессов и общей энергии жизнедеятельности связано с совершенствованием органов дыхания. У низших хордовых (многие оболочники, бесчерепные) этому служит увеличение размеров глотки и числа жаберных щелей.
Повышение уровня окислительных процессов и общей энергии жизнедеятельности связано с совершенствованием органов дыхания. У низших хордовых (многие оболочники, бесчерепные) этому служит увеличение размеров глотки и числа жаберных щелей.
Питание и пищеварение последовательно интенсифицируются в ряду хордовых животных. Усиление тока воды и образование слизистой сети в глотке у низших хордовых увеличило улавливание пищевых частиц. У позвоночных животных возник аппарат активного захвата пищи (челюсти, у некоторых групп — и конечности), а совершенствование зрения, обоняния и слуха позволило перейти к активным поискам и добыванию корма. Морфологическая и функциональная дифференцировка пищеварительного тракта привела к появлению в нем «конвейера ферментов»: определенные ферменты размещены в разных участках пищеварительной трубки, последовательно подвер
гая гидролизу пищевой комок. Это обеспечило специфические для каждого фермента условия и повысило скорость и эффективность пищеварения. Как выросты пищеварительного тракта возникают печень и поджелудочная железа, первая появилась уже у бесчерепных. В печени осуществляется синтез животного сахара — гликогена, креатина и мочевины, идет детоксикация (обезвреживание) чужеродных белков и разрушение старых эритроцитов; образующаяся в печени желчь поступает в кишечник и участвует в омылении (эмульгировании) жиров. В поджелудочной железе образуются пищеварительные ферменты, поступающие по протокам в начальную часть кишечника; она является и железой внутренней секреции, вырабатывающей гормоны инсулин и глюкагон.
Возникновение характерной для хордовых (кроме оболочников) замкнутой кровеносной системы, образование разделенного на камеры сердца и появление эндотелиальной выстилки стенок кровеносных сосудов усилило активную регуляцию и стабильность виутреиией среды организма. В эволюционном ряду хордовых животных росла устойчивость осмотического давления, содержания воды, солей, белков; заметно увеличивалось содержание сахаров в плазме крови. В повышении буфериости крови (ее способности сохранять определенный уровень pH) важную роль играет соотношение ионов одновалентных и двухвалентных металлов. В крови большинства беспозвоночных отно- Na 4- К
шение колеблется в пределах 1,5—7, тогда как у позвоноч
ных оно значительно выше (10—60). Повышение содержания углеводов в крови сопровождало увеличение подвижности; оио особенно возросло при переходе от экзотермных (пойкилотермных или холоднокровных) животных к эндотермным (гомойотермиым или теплокровным). Увеличение белков в плазме крови связано с повышением сопротивления инфекциям и усилением механизмов активного иммунитета.
В регуляции и стабилизации внутренней среды важную роль играют органы водного обмена и выделения. У бесчерепных функционирует выделительная система нефридиальноготипа, напоминающая аналогичную систему кольчатых червей. У позвоночных на той же основе возникают новые органы выделения — почки, выводящие из организма продукты распада и принимающие участие в водном и солевом обмене. В водном обмене и выделении участвуют также жабры, кожа и ее железы.
Усложнение строения организма хордовых сопровождалось усилением регуляции и координации процессов жизнедеятельности с помощью центральной нервной системы и функционально связанной с ней гормональной или химической регуляцией. По сравнению с беспозвоночными обе системы отличаются сложностью.
Нигде в животном иорстве не обнаружено такого разнообразия гормонов, работающих интегрированно, как в гормональной системе хордовых и особенно позвоночных (Баррингтон, 1964). Только у них встречаются тиреондные гормоны (иодированные аминокислоты — иодтиронины, тнреоглобулнны). У низших хордовых они выделяются эндостилем глотки, а у позвоночных — образующейся на его месте
щитовидной железой."Йодсодержащие гормоны (тироксин, трииодти- ронин) регулируют рост, развитие и метаморфоз, поддерживают нормальный уровень функционирования нервной системы и сердечной мышцы, регулируют потребление кислорода тканями, контролируют линьку роговых образований кожи. Гормоны щитовидной железы влияют на секреторную активность коры надпочечников. В свою очередь секреторная активность щитовидной железы находится под воздействием тиреотропного гормона гипофиза.
Стероидные гормоны, выделяемые корой надпочечников и интерстициальной тканью половых желез, регулируют водно-солевой и углеводный обмен, проницаемость клеточных мембран, стимулируют секреторную деятельность желез желудка, регулируют сезонную ритмику половых желез и половое поведение. Мозговое вещество надпочечников выделяет гормоны (адреналин, норадреналин), которые участвуют в передаче нервных импульсов, регулируют силу сокращений мышц сердца, кровеносных сосудов и кишечника, содержание сахаров в крови и тканях и многие другие процессы. Гормоны паращито- видных желез регулируют обмен кальция и фосфора. Островки Лан- герганса поджелудочной железы секретируют инсулин и глюкагон, воздействуя на углеводный обмен и содержание сахара в крови.
Особая роль принадлежит гормонам, выделяемым гипофизом. Они регулируют сезонную перестройку метаболизма, размножение, линьку, миграции, спячку. Тиреотропный, или тнреостимулирующий, гормон (ТСГ) стимулирует деятельность щитовидной железы. Гонадотропные гормоны (ГТГ) стимулируют образование и рост половых клеток и гормональную деятельность половых желез, усиливают действие гормонов, определяющих половое поведение, заботу о потомстве и лактацию (у млекопитающих). Антидиуретический гормон (АДГ) стимулирует секрецию гормонов надпочечниками, влияет на всасывание воды в почечных канальцах и на сокращения стенок капилляров. Гормоны роста, или соматотропные гормоны (СТГ), стимулируют рост, ускоряют синтез белков, способствуют росту костей, влияют на гормональную секрецию поджелудочной железы. Аденокортикотроп- ные гормоны (АКТГ) стимулируют секреторную активность коры надпочечников. Меланоцитостимулирующий гормон (МСГ) регулирует образование и распределение пигментов. Гормональная деятельность гипофиза регулируется центральной нервной системой (преимущественно гипоталамусом); на него влияют и гормоны других эндокринных желез.
Совокупность нервных и гормональных механизмов представляют пример кибернетической системы, построенной на отрицательных и положительных обратных связях. Эго обеспечивает постоянное приспособление организмов к меняющейся внешней среде и устойчивость их внутренней среды. Функциональная зависимость эндокринных желез друг от друга и регулирующая роль нервной системы создали у хордовых животных механизм, обеспечивающий подготовку и осуществление сезонных явлений: размножения, линьки, миграций, спячки и т. д. Сезонная перестройка обмена веществ и поведения обычно требует времени, растягиваясь на недели и месяцы, но обеспечи
вает состояние предварительной готовности к приближающимся переменам условий существования. Другие гормоны действуют быстрее. Так, поступление пищи в двенадцатиперстную кишку сразу же вызывает выделение гормонов, стимулирующих поступление в кишечник желчи и ферментов поджелудочной железы.
Неизмеримо быстрее работают механизмы нервной регуляции. Ответ на раздражение занимает доли секунды, что особенно необходимо высоко подвижным организмам, встречающимся со множеством быстро меняющихся условий, событий и явлений. Поэтому развитие подвижности в разных группах хордовых животных идет параллельно с совершенствованием центральной нервной системы и органов чувств.
Центральная нервная система бесчерепных имеет вид трубки. У оболочников трубчатая нервная система с расширением («головным мозгом») в передней части образуется у личинок, но в ходе метаморфоза редуцируется, частично сохраняясь в виде спинного ганглия. У позвоночных нервная трубка дифференцируется на головной и спинной мозг. Число нервных клеток головного мозга огромно и различно в разных классах; у млекопитающих их больше миллиарда. Вместе с рецепторами они создают сложные нейронные цепи, обеспечивающие сбор, передачу и переработку информации о состоянии организма н внешней среды, формируют и передают команды, по которым осуществляются ответные реакции организма.
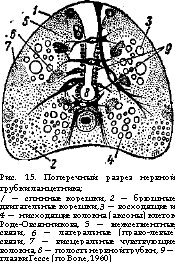
С развитием центральной нервной системы усложняются органы чувств. У низших хордовых они относительно просты. Рассеянные по поверхности тела чувствующие клетки, глазки Гессе (комплексы из пигментной и чувствующей клеток) в нервной трубке, статсцисты (органы равновесия оболочников) воспринимают механические, световые и химические сигналы из внешней среды. Чувствующие клетки имеются и во внутренних органах (интерорецепторы). У позвоночных животных сохраняются осязательные, термо- и хеморецепторы в коже, ротовой полости и т. д. Одновременно формируются специальные органы чувств, где наряду с нервными клетками — собственно рецепторами, развиваются так называемые сервомеханизмы — структуры, обеспечивающие отбор биологически важных сигналов и их усиление, что увеличивает возможность ориентации в окружающей среде. Таковы сложные камерные глаза, органы слуха, органы обоняния, боковой линии (у водных животных) и др Как правило, органы чувств парные, что отвечает принципу двухсторонней симметрии и увеличивает их разрешающую способность при ориентации в пространстве.
Еще по теме Основные черты организации хордовых:
- ГЛАВА 5 Основные черты и этапы истории жизни на Земле
- ГЛАВА 4 Организация жизни и ее основные характеристик
- 3.5.3. Проявление основных свойств материала наследственности и изменчивости на хромосомном уровне его организации
- ТИП ХОРДОВЫЕ — CHORDATA
- ТИП ХОРДОВЫ
- Общие черты, элементы систематики и распространение
- Глава 8. Гпавные черты процесса эволюции
- 13.5.4. Характеристика типа Хордовые
- Происхождение хордовых животных
- НЕКОТОРЫЕ ЧЕРТЫ БИОГЕОГРАФИЧЕСКОГО ДЕЛЕНИЯ МИРОВОГО ОКЕАНА
- НИЗШИЕ ХОРДОВЫЕ, ИХ ПРОИСХОЖДЕНИЕ И ЭВОЛЮЦИЯ
- Значение хордовых животных для человека
- Общие черты в строении и развитии ленточных червей в связи с их паразитическим образом жизни
- 7.3. МОРФОФИЗИОЛОГИЧЕСКИЕ И ЭВОЛЮЦИОННЫЕ ОСОБЕННОСТИ ЯИЦ ХОРДОВЫХ
- ОТКУДА ВЗЯЛИСЬ ХОРДОВЫЕ
- СТРОЕНИЕ ХОРДОВОГО ЖИВОТНОГО НА ПРИМЕРЕ РЫБЫ
- ГЛАВА 14 ФИЛОГЕНЕЗ СИСТЕМ ОРГАНОВ ХОРДОВЫХ
- Н. П. НАУМОВ, Н. Н. КАРТАШЕВ. зоология позвоночных ЧАСТЬ 1 НИЗШИЕ ХОРДОВЫЕ, БЕСЧЕЛЮСТНЫЕ, РЫБЫ, ЗЕМНОВОДНЫЕ, 1979