Кровеносная система и кровообращение
Кровеносные системы костных и хрящевых рыб сходны; различия сводятся лишь к некоторым деталям (рис. 120). Сердце двухкамерное (одно предсердие + один желудочек). У всех костных рыб есть венозный синус или венозная пазуха (sinus venosus), из которого кровь поступает в предсердие (atrium) и из него — в желудочек (ventriculus).
У кистеперых, двоякодышащих, осетрообразных, многоперов и панцирникообразных хорошо развит артериальный конус (conus arteriosus), как и у хрящевых рыб, несущий на внутренних стенках полулунные клапаны. У амии артериальный конус сильно редуцирован, а у остальных костных рыб (Teleostei:) от артериального конуса остаются лишь один-два ряда клапанов перед входом в брюшную аорту. При редукции артериального конуса образуется толстостенное вздутие начальной части брюшной аорты — луковица аорты (bulbus aortae), имеющая стенки с гладкой мускулатурой и смягчающая пульсовые толчки. Кровяное давление у большинства костных рыб несколько выше (18—120 мм рт. ст.), чем у хрящевых (7—45 мм).
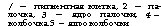 Брюшная аорта (aorta ventralis) распадается на 4 пары приносящих жаберных артерий (рис. 120, 6), снабжающих кровью расположенные на I—IV жаберных дугах жабры. Окислившаяся в капиллярах жаберных лепестков кровь собирается в выносящие жаберные артерии (их тоже 4 пары), которые впадают в корни спинной аорты (рис. 120, 8), позади жаберного отдела сливающиеся в спинную аорту (aorta dorsalis) (рис. 120,
Брюшная аорта (aorta ventralis) распадается на 4 пары приносящих жаберных артерий (рис. 120, 6), снабжающих кровью расположенные на I—IV жаберных дугах жабры. Окислившаяся в капиллярах жаберных лепестков кровь собирается в выносящие жаберные артерии (их тоже 4 пары), которые впадают в корни спинной аорты (рис. 120, 8), позади жаберного отдела сливающиеся в спинную аорту (aorta dorsalis) (рис. 120,
11). Спереди корни аорты, отделив сонные артерии (рис 120, 10), тоже соединяются друг с другом (рис. 120,
 9), образуя характерный для костных рыб головной круг. Лежащая под позвоночным столбом спинная аорта последовательно ответвляет артерии, снабжающие кровью мускулатуру и внутренние органы, и уходит в гемаль- ный канал хвостового отдела позвоночного столба; здесь ее называют хвостовой артерией (a.
9), образуя характерный для костных рыб головной круг. Лежащая под позвоночным столбом спинная аорта последовательно ответвляет артерии, снабжающие кровью мускулатуру и внутренние органы, и уходит в гемаль- ный канал хвостового отдела позвоночного столба; здесь ее называют хвостовой артерией (a.
 Собравшая венозную кровь из хвостового стебля хвостовая вена (vena caudalis) (рис. 120, 16) входит в полость тела и разделяется на правую и левую воротные вены почек (v. portae renalis) (рис. 120, 17). У части костных рыб, как и у хрящевых, они входят в почки и распадаются там на капилляры, т. е. образуют воротную систему почек; почечные капилляры, сливаясь, впадают в задние кардинальные вены. Однако у многих костистых рыб в правой стема частично редуцируется: часть крови переходит в правую заднюю кардинальную вену, минуя капилляры. Несущие кровь из головы передние кардинальные вены (v. cardinalis
Собравшая венозную кровь из хвостового стебля хвостовая вена (vena caudalis) (рис. 120, 16) входит в полость тела и разделяется на правую и левую воротные вены почек (v. portae renalis) (рис. 120, 17). У части костных рыб, как и у хрящевых, они входят в почки и распадаются там на капилляры, т. е. образуют воротную систему почек; почечные капилляры, сливаясь, впадают в задние кардинальные вены. Однако у многих костистых рыб в правой стема частично редуцируется: часть крови переходит в правую заднюю кардинальную вену, минуя капилляры. Несущие кровь из головы передние кардинальные вены (v. cardinalis
anterior) (рис. 120, 19), собирающие кровь из грудных плавников подключичные вены (v. subclavia) (рис. 120, 20) и задние кардинальные вены (v. cardinalis posterior) (рис. 120, 18) каждой стороны сливаются в соответствующий — левый или правый кювьеров проток (ductus cuvieri), впадающие в венозную пазуху. Вены, собирающие кровь от желудка, селезенки и всего кишечника, сливаются в воротную вену печени (v. portae hepatis); она входит в печень и распадается на капилляры, образуя воротную систему печени. Печеночные капилляры вновь сливаются в вены, объединяющиеся в короткую печеночную вену (v. hepatica) (рис. 120, 24), впадающую непосредственно в венозную пазуху.
В разных группах костных рыб встречаются отклонения от описанной схемы. Наиболее своеобразны особенности кровеносной системы двоякодышащих рыб, обусловленные наличием у них добавочного легочного дыхания; с ним связано возникновение легочного круга кровообращения. Предсердие свисающей со спинной стороны перегородкой (рис. 121, 5) почти полностью разделяется на две половины: в левую впадает несущая кровь из легких — легочная вена, а в правую открывается венозная пазуха. Эта перегородка в виде складки проходит через желудочек и продолжается в спиральный клапан артериального конуса. От брюшной аорты отходят 5 пар приносящих жаберных артерий (передняя из них проходит по подъязычной дуге).
Через капиллярные сети жабр приносящие артерии переходят в выносящие жаберные артерии, впадающие в корни аорты; последние продолжаются вперед как сонные артерии, а позади жаберной области сливаются в спинную аорту. От последней (пятой) выносящей жаберной артерии отходит легочная артерия (рис. 121, 12), ветвящаяся в легком. При достаточном содержании кислорода в
Рис. 121. Схема жаберно-легочного кровообращения двоякодышащих рыб (по Гудричу, упрощено):
/ — венозная пазуха, 2 — предсердие, 3 — желудочек. 4 — артериальный конус, 5 — перегородка предсердия, переходящая в складку верхней стенки желудочка н продолжающаяся в спиральный клапан артериального конуса. 6 — брюшная аорта, 7 — приносящие жаберные артернн, 8 — выносящие жаберные артернн, 9 — кории аорты, 10 — спиииая аорта, // — сониая артерия, 12 — легочная артерия, 13 — легкое, 14 — легочная вей а, 15 — кювьеров проток, 16 — задняя полая веиа; /—V жаберные щели
воде, когда полное насыщение крови кислородом Происходит в жабрах, легкое не функционирует и получает артериальную кровь.
При неполном насыщении крови кислородом в жабрах в легкое поступает лишь частично окисленная кровь и там доокисляется. В правую половину предсердия поступает венозная кровь из венозной пазухи, в левую артериальная кровь из легкого. Перегородка предсердия затрудняет перемешивание крови. При сокращении предсердия более артериальная кровь течет вдоль складки желудочка и спирального клапана артериального конуса и попадает преимущественно в первые приносящие жаберные артерии, получает дополнительно кислород в жабрах и идет в голову и корни аорты. Более венозная кровь из правой части предсердия идет преимущественно в задние приносящие жаберные артерии, получает в жабрах какое-то количество кислорода и попадает в корни аорты, а частично — через легочную артерию — в легкие. Большой и малый круги кровообращения, таким образом, разобщены очень слабо. Однако это делает возможным одновременное использование жаберного и легочного дыхания; причем более артериальная кровь поступает в голову, а по телу и в легкие идет смешанная кровь. Есть своеобразные черты и в периферической системе. Задние кардинальные вены развиваются относительно слабо. Основная масса крови из капилляров обеих почек в конечном счете попадает в крупный сосуд — заднюю полую вену (v. cava posterior), впадающую непосредственно в венозную пазуху (рис. 121, 16).
Таким образом, у двоякодышащих появляются изменения, которые напоминают особенности кровеносной системы земноводных. Можно предполагать, что у имевших легкие кистеперых — предков земноводных — кровеносная система была сходна с кровеносной системой ныне живущих двоякодышащих. Поэтому изучение становления и функционирования кровеносной системы двоякодышащих позволяет представить возможные пути морфологических перестроек, сопровождавших обособление первого класса наземных позвоночных — земноводных — от рыбообразных предков.
Основные показатели крови (кислородная емкость, количество гемоглобина и эритроцитов, ионный состав) обычно заметно выше у более подвижных пелагических видов и меньше — у малоподвижных донных форм. В пределах класса костных рыб общее количество крови варьирует от 1,1 до 7,3% от массы тела, число эритроцитов — от 580 тыс. до 4,1 млн, в 1 мм8, содержание гемоглобина изменяется в пределах 1,1—17,4 г% и 0,5—3,4 г на 1 кг массы; кислородная емкость крови составляет 1,5—23,0 объемных процента. Верхняя граница этих показателей у костных рыб заметно выше, чем у хрящевых рыб (см. табл. 5, с. 156). Живущие в водах Антарктиды белокровные рыбы (около 10 видов сем. Chaenichthyidae, подотр. нототениевые, отр. окунеобразные) в отличие от других рыб не имеют эритроцитов и гемоглобина. Кровь у них бесцветная. Органами дыхания служат не столько жабры, сколько обильно снабженная капиллярами кожа (до 45 мм длины капилляров на 1 мм2 поверхности тела). Поверхность капилляров кожи и плавников в 2 раза превышает общую поверхность тела. Живут эти рыбы в воде с высоким содержанием кислорода и
постоянно низкой температурой (ниже +2° С). Отличаются большими размерами сердца и низким содержанием кислорода в крови.
Важную роль в обеспечении энергетики организма играют углеводы, циркулирующие в крови (глюкоза, гликоген и др.). В отличие от высших позвоночных у рыб колебания сахара в крови очень велики, что связано с несовершенным механизмом регуляции. Выявляются сезонные изменения: повышение сахаров в периоды размножения и миграций (обычно в теплое время года) и снижение на зимовках, при малоподвижном образе жизни. Регуляция обеспечивается использованием запасов гликогена в печени под контролем гормональной и нервной систем, т. е. обычным путем сезонных перестроек метаболизма. У высокоподвижных и активных рыб (тунцы и др.) содержание сахара в крови удерживается в пределах 60—90 мг% (у осетровых поднимается до 115 мг%), а у малоподвижных донных рыб (морской чёрт) обычно не бывает выше 10 мг% (у хрящевых — 20—50 мг%). Кровь костных рыб отличается от крови хрящевых рыб большим содержанием белков в плазме. Образование форменных элементов крови происходит преимущественно в селезенке и в почках (особенно в их передних отделах).
Органы выделения и водно-солевой обмен. Система органов выделения, основу которой у всех рыб составляют почки, не только выводит продукты распада азотистых веществ, но и обеспечивает физикохимическую устойчивость внутренней среды организма: осмотического давления, кислотно-щелочного ионного равновесия. В этих процессах участвуют почки, жаберный аппарат, кожа, пищеварительная трубка и печень. Эти системы различаются у хрящевых и костных рыб.
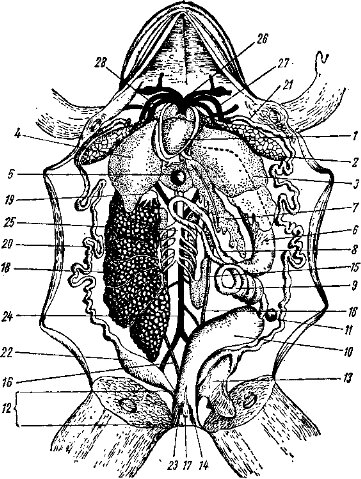
Парные мезонефрические (туловищные) почки костных рыб лежат ниЖе позвоночного столба, почти вдоль всей полости тела. У многих видов в задней половине правая и левая почки сливаются вместе. Вольфовы каналы выполняют функцию мочеточников. По выходе из почки мочеточники правой и левой почек сливаются и, образовав тонкостенный полый вырост — мочевой пузырь (vesica urinaria) (рис. 122, 3), открываются наружу мочевым отверстием (у двоякодышащих — в клоаку). Почки получают артериальную кровь из почечных артерий; большие порции венозной крови приходят сюда по воротным венам почек из хвостового отдела. У пресноводных рыб хорошо развиты боуменовы капсулы с крупными клубочками. У морских костных рыб размеры клубочков уменьшаются, а у нескольких десятков видов из разных отрядов (жабы-рыбы, некоторые морские иглы и др.) клубочки практически редуцированы (агломерулярные почки).
У подавляющего большинства лучеперых рыб, особенно у костистых, конечным продуктом распада азотистых (в том числе и белковых) соединений, выводимым из организма, служит аммиак (как и у большинства беспозвоночных). Это парадоксальное явление находит свое объяснение в том, что жаберные лепестки этих рыб способны интенсивно выделять в окружающую среду аммиак и ионы солей, обеспечивая таким образом значительную часть процессов выведения продуктов распада из организма. Это объясняет, почему у многих видов почки
)
Рис. 122. Выводные каналы мочеполовой системы самца щуки —
Esox lucius (по Гуртовому):
/ _ почка, 2 — мочеточник, 3 — мочевой пузырь, 4 — наружное мочевое отверстие, 5 — семенник, б — семяпровод, 7 — наружное полозое отверстие, 8 — кишечник, 9 — анальное отверстие ц
(особенно их верхние части) потеряли исходную функцию, превратившись в лимфоидный орган, участвующий в кроветворении. Часть азота выводится в виде мочевины, триметиламиноксида, креатина и близких соединений; она возрастает у морских видов по сравнению с пресноводными. Двоякодышащие рыбы, впадающие в оцепенение при высыхании водоемов (протоптерус), в активном состоянии выделяют аммиак, а в оцепенении — мочевину, накапливающуюся в организме. Она выводится после пробуждения рыбы (напомним: аммиак много токсичнее мочевины.)
Выделение продуктов распада у рыб тесно связано с водно-солевым обменом, обеспечивающим осмотическое и кислотно-щелочное равновесие в тканях. У морских и пресноводных рыб эти процессы протекают различно. Пресноводные рыбы живут в гипотонической среде (осмотическое давление в их тканях значительно выше осмотического давления пресной воды (табл. 6, с. 158). Поэтому вода постоянно проникает в организм через кожу (рис. 123, /), жабры и с пищей. По некоторым данным за 3—5 ч поступающей извне водой замеряется вся вода организма; ему постоянно грозит излишнее «обводнение». Высоко развитый фильтрационный аппарат почек позволяет выводить избытки воды; за сутки пресноводные рыбы выделяют от 50 до 300 мл мочи и более на 1 кг массы тела. Потери солей при выведении больших количеств мочи компенсируются активной реабсорбцией солей в почечных канальцах и поглощением солей жабрами из внешней среды;
часть солей поступает с пищей. Такой пресноводный тип водно-солевого обмена возник у предков рыб в пресных водоемах и сохранился у современных пресноводных костных рыб.
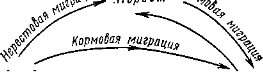




 При переходе в море костные рыбы оказались в резко гипертонической среде, грозившей им «иссушением» вследствие потерь воды через кожу и жабры, с мочой и фекалиями (рис. 123, II); некоторое повышение осмотического давления в крови и тканях по сравнению с пресноводными формами (табл. 6, с. 158) предотвратить эти потери не могли. Возник новый механизм регуляции водно-солевого обмена: уменьшилось число клубочков в почках, а у части видов они исчезли полностью (агломерулярные почки); эти рыбы выделяют лишь 0,5—20мл воды на 1 кг массы в сутки. Взамен усиливается роль почечных канальцев, железистые клетки стенок которых увеличили выделение мочевины и других продуктов азотистого обмена. Выделение солей жабрами обеспечено особыми клетками жаберных лепестков. На этой базе водный баланс морских костных рыб был восстановлен тем, что рыбы стали пить морскую воду (от 40 до 200 мл на 1 кг массы в сутки), а избытки солей, поступавшие при этом в организм, выделяли через жабры и с каловыми массами.
При переходе в море костные рыбы оказались в резко гипертонической среде, грозившей им «иссушением» вследствие потерь воды через кожу и жабры, с мочой и фекалиями (рис. 123, II); некоторое повышение осмотического давления в крови и тканях по сравнению с пресноводными формами (табл. 6, с. 158) предотвратить эти потери не могли. Возник новый механизм регуляции водно-солевого обмена: уменьшилось число клубочков в почках, а у части видов они исчезли полностью (агломерулярные почки); эти рыбы выделяют лишь 0,5—20мл воды на 1 кг массы в сутки. Взамен усиливается роль почечных канальцев, железистые клетки стенок которых увеличили выделение мочевины и других продуктов азотистого обмена. Выделение солей жабрами обеспечено особыми клетками жаберных лепестков. На этой базе водный баланс морских костных рыб был восстановлен тем, что рыбы стали пить морскую воду (от 40 до 200 мл на 1 кг массы в сутки), а избытки солей, поступавшие при этом в организм, выделяли через жабры и с каловыми массами.
У проходных рыб при переходе из моря в реки и наоборот происходит перестройка водно-солевого обмена. Так, например, у речного угря — A. anguilla:
Выделяется мочи
мл/кг/сут Д мочн Д крови
В реке (Д среды—0,06) 60—150 0,09 0,63
В море (Д среды—1,85) 2—4 0,79 0,82
Огшсанные адаптации водно-солевого обмена позволили костистым рыбам широко освоить пресные и соленые водоемы. Напомним, что хрящевые рыбы, удерживая в организме мочевину, эффективно адаптировались к жизни в морях, но практически не освоили пресные воды.
Половая система и особенности размножения. Половые железы (яичники у самок и семенники у самцов) у костных рыб парные, висят на брыжейке под почками, по бокам плавательного пузыря. Взаимоотношения половых и мочевых протоков в разных группах варьируют. У самцов низших костных рыб (двоякодышащие, надотряд ганоидные) уменьшается связь семенников с почкой: семявыносящие канальцы впадают в небольшое число каналов задней (а не передней,
как у хрящевых рыб) части почки или лишь в один (Frotopterus), которые открываются в вольфов канал, на выходе из почки функционирующий как семяпровод и мочеточник. У самцов костистых рыб вольфовы каналы несут только функцию мочеточников (см. рис. 122, 2), а канальцы семенника открываются в семяпровод — новообразование, не связанное с протоками мезонефроса. В нижней части семяпровода иногда образуется расширение — семенной пузырек. Семяпроводы обоих семенников открываются наружу общим половым (рис. 122, 7) или мочеполовым отверстием.
У самок двоякодышащих, как и у хрящевых рыб, длинные яйцеводы —- мюллеровы каналы — открываются в полость тела воронками, а задними концами — в клоаку. У самок осетрообразных \ многоперообразных и амиеобразных короткие и широкие яйцеводы — мюллеровы каналы — воронкой открываются в полость тела около яичников, а задним концом — в мочеполовой синус. У самок панцирни- кообразных и у подавляющего большинства костистых рыб мюллеровы каналы редуцируются; оболочка мешкообразного яичника продолжается в виде короткого протока, открывающегося либо в мочеполовой синус, либо в самостоятельное половое отверстие. Таким образом, в отличие от всех других групп рыб у них зрелое яйцо не выпадает в полость тела, а попадает в полость яичника и по короткому протоку выбрасывается наружу. Лишь у лососевых и немногих других костистых рыб (клюворылообразные, муреновые из угреобразных и др.) зрелые яйца выпадают из яичника в полость тела и выводятся наружу через половые поры или очень короткие яйцеводы с широкой воронкой.
Большинство видов костных рыб раздельнополы. Среди каменных окуней — Serranidae и морских карасей — Sparidae (окунеобразные) известен нормальный гермафродитизм; каждая особь имеет и мужскую, и женскую половые железы, обычно созревающие поочередно, что предотвращает самооплодотворение. У некоторых окунеобразных (Labroides и др.) обе железы могут функционировать параллельно, что сопровождается быстрым — за несколько минут — превращением особи из самца в самку и обратно (рис. 124). При отсутствии партнера одиночная рыба сначала откладывает икру, а затем поливает ее спер- миями. У морской Onigocia macrolepis (Platycephalidae) происходит реверсия (обращение) пола. Половые железы у молодых рыбок развиваются в виде двухполой гонады, в которой вентральная часть представляет семенник, а дорзальная — яичник. Созревшие особи вначале функционируют как самцы, а во вторую половину жизни как самки. При этом «семенниковая» часть деградирует, а яичник разрастается (Такеда, 1970).
Оплодотворение наружное, в воде; у немногих видов оно внутреннее. При этом семя вводится в половые пути самки при помощи сильно развитого генитального сосочка (бычки подкаменщики — Cottidae и др.) или при помощи гоноподия — измененного участка анального
1 Мюллеровы каналы сохраняются и у самцов осетрообразных рыб. Фуикцня их не ясна; возможно, они служат для удаления наружу избытка целомической жидкости.
плавника (многие карпозубообразные; рис. 125). У фаллостето- видных (из атеринообразных) у самцов на нижней части головы расположен своеобразный совокупительный орган — приапий (см. рис. 103).
 Время наступления половой зрелости различно у разных видов и варьирует в пределах одного вида. Обилие корма, повышенные температуры способствуют более быстрому росту и ускоренному половому созреванию. Поэтому, например, плотва R. rutilus в Финляндии становится половозрелой в возрасте 5—6 лет, а в Южной Европе — в 3 года. Мелкие карпозубые созревают в возрасте 2—4 месяцев, кильки каспийские — Clupeonella delicatula, хамса — Engraulis encrasicholus — в годовалом возрасте, щука — Esox lucius —в 4 года (в некоторых водоемах даже в 2), сазан —
Время наступления половой зрелости различно у разных видов и варьирует в пределах одного вида. Обилие корма, повышенные температуры способствуют более быстрому росту и ускоренному половому созреванию. Поэтому, например, плотва R. rutilus в Финляндии становится половозрелой в возрасте 5—6 лет, а в Южной Европе — в 3 года. Мелкие карпозубые созревают в возрасте 2—4 месяцев, кильки каспийские — Clupeonella delicatula, хамса — Engraulis encrasicholus — в годовалом возрасте, щука — Esox lucius —в 4 года (в некоторых водоемах даже в 2), сазан —
РИС‘ 12о\унРя?™Йо^Х?/ОДИТИЗМ СУРНпт CClrPi0 ~ В 2—4 гОДа-
белуга — Н. huso — в 9—23, а
Раба с полосами на теле (самцовая фаза) в я ^ А „ лил.
паре с самкой (без полос) поднимаются вверх СвВрЮГа — ACipetlSer StellCltUS —
и выпрыгивают из воды» самка вьгметьгвает р Q оо г/’мтя Г'яитти лйипип
икру, а самец ее оплодотворяет (вверху). Затем 0 Й ^ Г0Даgt; ^аМЦЫ ООЫЧНО
оба заиыривают (средний рисунок), переходя СОЗреваЮТ раньше СаМОК. У боЛЬ-
в противоположную фазу («самец в самку» . . _
н, наоборот, «самка в самца») (виизу). Циклы ШИНСТВЗ СамКИ НвСКОЛЬКО КруП-
повкфяются несколько раэ ^ нее самЦОВ, а у ВИДОВ, у КО-
торых самцы охраняют кладку, они, наоборот, немного крупнее самок. У части видов самцы отличаются от самок по окраске, по длине и форме плавников. У немногих видов к началу периода размножения развивается брачный наряд: становится более яркой окраска, может даже изменяться форма тела, как у некоторых лососей. У некоторых глубоководных рыб, живущих рассредоточенно, когда встречи полов затруднены и не всегда гарантированы, имеются карликовые самцы, в течение всей жизни прикрепленные к самкам.
Плодовитость рыб в среднем много выше плодовитости наземных позвоночных, что определяется высокой смертностью (особенно гибелью икры и мальков) от хищников и других факторов (обсыхания, занесения илом и др.). Молодые самки обычно откладывают меньше икры, чем самки старшего возраста, виды более крупных размеров чаще более плодовиты, чем мелкие. При обилии пищи плодовитость выше, чем при ее недостатке. Наиболее плодовиты некоторые виды, отклады-
Рис. 125. Гамбузия — Gambusia afjinis. Самка (вверху), самец и его гоноподий (внизу)
вающие плавающую (пелагическую) икру. Луна-рыба — М. mola откладывает до 300 млн. икринок (в течение года выживает менее 1% молоди), треска — Gadus morhua — 2,5—10 млн., палтус — Н. hippoglossus — 2—3,5 млн. В то же время у откладывающих пелагическую икру анчоусов и килек плодовитость много ниже — всего 10—60 тыс. икринок. Среди наших пресноводных рыб наиболее плодовиты сазан (до 1,5 млн.), щука (100 тыс. — 1 млн.), судак (200 тыс.— млн.), лещ (90—350 тыс.). Количество выметываемых сперматозоидов много выше. Так, у леща массой 250 г выделяется до 150 биллионов сперматозоидов.
Пелагическая, т. е. свободно плавающая в толще воды икра обычно мелкая, с довольно большой жировой каплей, увеличивающей ее плавучесть. У других видов плавучесть обеспечивается либо обзодне- нием наружной оболочки, либо образованием на ней выростов. У видов, нерестящихся на дне или среди зарослей подводной растительности, икра демерсальная — крупная и тяжелая; наружная оболочка у многих видов в воде становится клейкой и прилипает к подводным предметам. У некоторых рыб оболочки отдельных икринок слипаются друг с другом, и кладка в виде лент развешивается на водорослях.
В разных группах костных рыб в той или иной степени проявляется забота о потомстве, заметно снижающая гибель икры и смертность молоди. Плодовитость таких рыб обычно сокращается. Лососевые рыбы закапывают икру в галечный грунт и несколько дней охраняют гнездо; кета выметывает 2—4 тыс. икринок, горбуша — 1—2 тыс.
alt="" />
Рис. 126. Забота о потомстве у костистых рыб:
А — самец пинагора — Cyclopterus lumpus (скорпенообразные) охраняет икру;
В — самец трехиглой колюшки — Gasterosteus aculeatus (колюшкообразные) охраняет гнездо с икрой; В — самец морской иглы — Syngnaihus typhle, вынашивающий икру в сумке (вверху), вскрытая сумка (внизу); Г — самка горчака — Rhodeus sericeus (карпообразные) откладывает икру в мантийную полость беззубки;
Д — самка сома Aspredo со вдавленными в кожу икринками, Е — самец Apogon (окунеобразные) вынашивает икру во рту (вверху — икринки на разных стадиях
развития)
Число скатившихся в море мальков составляет около 2—3% от числа выметанных икринок (с. 188). Устраивают примитивное гнездо из растений, камешков или роют ямки и охраняют отложенную икру некоторые сомы, ильная рыба, двоякодышащие — Protopterus и Lepidosiren, многие губановые и бычковые, из окунеобразных и др. Самцы колюшек из обрывков растений, скрепляемых загустевающей в
воде слизью, строят полое шаровидное гнездо (с. 201; рис. 126) и охраняют его. Самец пинагора (скор пенообразные) при помощи брюшной присоски удерживается около отложенной на мелководье кучки икры (рис. 126), охраняет ее и во время отлива смачивает водой; вылупившаяся молодь первое время присосками прикрепляется к самцу. Некоторые виды мелких тропических пресноводных рыб устраивают на поверхности воды гнездо из пузырьков воздуха, удерживаемых загустевающей слизью; отложенные в гнездо икринки охраняются самцом (лабиринтовые рыбы — гурами, петушки, макроподы). У самок горчаков образуется длинный яйцеклад, при помощи которого небольшая порция икры откладывается в мантийную полость двустворчатых моллюсков, где и оплодотворяется проникшими с током воды сперматозоидами. Вылупившиеся из икринок личинки некоторое время остаются здесь же, под защитой раковины, удерживаясь в складках мантии при помощи крючковидных выростов желточного мешка и колючих чешуек.
В разных группах рыб икра вынашивается в ротовой полости (некоторые мелкие сомовые, цихловые из окунеобразных и др.). У сомов икру вынашивает самец, у тиляпий (цихловые) — самка, у апагоновых — оба пола. Икра полностью заполняет ротовую полость (рис. 126), и рыба в этот период не питается. Вылупившаяся молодь некоторое время при опасности прячется во рту взрослой рыбы. Самка тиляпии держится около выводка и при опасности, совершая зигзагообразные движения, увлекает его за собой. В основе этого явления лежит свойственный молоди рефлекс следования (рис. 127). Самка южноамериканского сома Aspredo выметывает икру на дно и после оплодотворения ложится на нее, вдавливая икринки в набухшую кожу брюха (см. рис 126). Через подходящие к каждой икринке кровеносные сосуды развивающиеся зародыши получают не только кислород, но и питательные вещества. У южноамериканского сома Tachysurus икра заглатывается и развивается в желудке; естественно, что в этот период рыба не питается, а ее пищеварительные железы временно не функционируют.
У самцов морских игл и коньков (колюшкообразные) на нижней стороне тела развивается образованная складками выводковая сумка (см. рис. 126), в которую самка откладывает несколько десятков икринок. Сеть капилляров на внутренней стенке сумки обеспечивает снабжение развивающихся зародышей кислородом. Молодь первое время держится рядом с самцом, при опасности скрываясь в сумке. Эти приспособления уменьшают смертность и улучшают условия развития зародышей. Они особенно распространены среди тропических видов, живущих в водоемах со сложными биоценозами, где каждый вид имеет много врагов, и часто неблагоприятен и кислородный режим.
У небольшого числа видов костистых рыб из разных групп развивается яйцеживорождение: икра задерживается в полости яичника или яйцевода и личинка вылупляется в момент икрометания (эндемики Байкала: голомянковые — Comephoridae из скорпенообразных; самка выметывает 800—2500 икринок) или личинки задерживаются в половых протоках до тех пор, пока у них полностью или частично не

JS**"

Рис. 127. Характер движения хемихромиса-красавца, вызывающего у мальков реакцию следовании (Л); сбор мальков самкой тиляпии (Б)
рассосется желточный мешок (гамбузии, меченосцы, молинезии и другие из карпозубообразных, часть бельдюговых — Zoarcoidae из окунеобразных, морские окуни — Sebastes из скор пенообразных и др.). Плодовитость яйцеживородящих рыб обычно невелика. Так, у европейской бельдюги — Zoarces viviparus длиной 25—30 см рождается от 10 до 400 мальков длиной 3,5—4 см. Лишь у морских окуней
сохраняется высокая плодовитость: самки разных видов выметывают до 10—350 тыс. мелких личинок длиной 4—6 мм.
Относительно немногие виды (тихоокеанские лососи, речной угорь,' байкальские голомянки, сельдь черноспинка и др.) относятся к моно- циклическим видам: все или значительная часть особей размножаются один раз в жизнн н после нереста гибнут. Большинство видов поли- цикличны: размножаются несколько раз в жизни. Численность таких видов в природе обычно более устойчива, чем моноциклических. У большинства рыб размножение приурочено к определенному периоду года, у некоторых тропических рыб в году может быть 2—3 цикл? размножения. Крупные осетровые и, видимо, очень крупные виды других отрядов размножаются раз в 3—5 лет. В умеренных и северных широтах размножение обычно проходит весной или в начале лета (большинство наших видов). Однако у многих лососевых нерест идет осенью; при длительном инкубационном периоде (до 100—180 дней) выклев личинок идет ранней весной. Налим нерестится зимой, в декабре—январе; выклевываются личинки через 1,5—2,5 месяца.
К началу размножения изменяется физиологическое состояние рыб, зависящее от условий жнзни в нагульный период (обеспеченности пищей, температуры и т. п.) и сказывающееся на размерах кладок. Появление обильных нли малочисленных кладок, успешность эмбриогенеза, степень выживания личинок обусловливают появление то «мощных», то «слабых» поколений, особенно отчетливо выраженное у плодовитых видов. Так, исландской трески рождения 1922 г. выловлено за ряд лет 705 тыс. т, а поколения 1927 г. — всего 19 тыс. т.
Для успешного размножения необходима встреча половозрелых и готовых к размножению самцов и самок на местах нереста. У большинства видов это обеспечивается образованием нерестовых стай, объединяющих особей разного пола и сходного физиологического состояния. У жнвущнх одиночно рыб встреча партнеров и синхрони зацня их готовности к размножению обеспечивается разнообразными нерестовыми сигналами — звуковыми, оптнческнмн, химическими, а у электрических рыб — и электрическими. Как говорилось выше, у некоторых малоподвижных и одиночно живущих глубоководных рыб (удильщики — Ceratioidei) маленькие самцы паразитируют на самках, превращаясь, по сути, в придаток самки, вырабатывающий сперму. Наличие таких карликовых паразитических самцов обеспечивает размножение малочисленных,-ведущих одиночный обра^ жизни видов. Как дополнительную гарантию размножения можно рассматривать и гермафродитизм (см. с. 235).
Осуществление нереста связано с наличием так называемых «нерестовых сигналов», непосредственно стнмулнрующих откладку икры и ее оплодотворение. К ним относятся ландшафтные особенности нерестилища (определенные грунты, наличие характерной водной растительности, температура, аэрированность воды и т. п.), состояние партнеров, готовых к размножению, в частности — их окраска и поведение. Существенную роль в синхронизации процессов нереста играют выделяемые в воду зрелыми самцами вещества — стимуляторы, осо
бенно стерогормоны копулины. Они ускоряют готовность самок к выметыванию икры или к осеменению (у живородящих видов).
Недостаток мест, пригодных для нереста, может ограничивать размножение и рост численности многих видов. Зарегулирование стока и замедление течения при строительстве плотин гидроэлектростанций нередко ухудшают возможности размножения видов, нерестующих на песчаных или галечных перекатах, а слабое развитие водной растительности в молодых водохранилищах ограничивает возможности размножения рыб, выметывающих икру на растительность. Поэтому при рациональном рыбном хозяйстве необходимо уделять внимание устройству искусственных нерестилищ и мелиорации (улучшению) естественных. Недостаток естественных нерестилищ у некоторых проходных рыб компенсируется существованием «озимых» и «яровых» рас (некоторые осетровые, лососевые, карповые). Озимая раса входит в низовья рек осенью с еще незрелыми половыми продуктами, зимует там и весной, рано приходя на нерестилища, выметывает икру. Яровая f аса входит в реки летом со зрелыми половыми продуктами, прямо идет на нерестилище и приступает к нересту позже озимой расы. Это дает возможность разным популяциям одного вида последовательно использовать одни и те же нерестилища. У части видов разделение на озимые и яровые расы позволяет увеличить количество используемых нерестилищ; озимая раса может дальше подняться вверх по реке и достигнуть нерестилищ в верховьях. Так, в р. Печору заходит преимущественно озимая раса семги, нерестящаяся в верховьях, а в мелкие речки Кольского полуострова, где подходящие для нереста места расположены недалеко от устья, — преимущественно яровая раса.
У большинства рыб с размножением связаны нерестовые миграции — переход с места нагула на места размножения. Нерестовые миграции составляют часть годового биологического цикла; степень их выражения у разных видов различна. У многих пресноводных (щука, окунь, пескари и пр.) и оседлых морских рыб (бычки, коралловые рыбы и др.) протяженность нерестовых миграций невелика и измеряется сотнями метров или немйогими километрами. Некоторые морские рыбы совершают кочевки в сотни и тысячи километров (сельди, тресковые, морские окуни и др.). Так, норвежская сельдь нерестует у берегов южной Норвегии и уходит для нагула к северу, возвращаясь на нерест в следующем году (рис. 128). Вылупившаяся из икры молодь Атлантическим течением уносится на северо-восток. Еще незрелые сельди начинают медленно двигаться к югу, ежегодно приближаясь к берегам и отходя в открытое море. Достигнув ко времени подхода к нерестовому району половой зрелости (в возрасте 5 лет), они нерестятся и уходят для нагула на север и восток, вновь возращаясь сюда лишь на следующий год. Миграции проходных рыб, идущих для икрометания из морей в реки (осетровые, лососевые) или из рек в море (речной угорь), имеют протяженность в сотни и тысячи километров.
Нерестовые миграции облегчают встречу половозрелых особей и обеспечивают возможность выметывания икры и развитие личинок и мальков в наиболее благоприятных условиях, повышающих их выживаемость. Места нереста многих видов рыб отличаются от мест

Рис. 128. Схема миграции норвежской сельди — Clupea harengus у берегов Южной Норвегии. Место нереста заштриховано:
О — Лофтенские о-ва (куда течением относит личинку). Молодь сельди: I — однолетки, II — двухлетки, III — трехлетки, IV — четырехлетки, V — сельди, достигшие половой зрелости
(нерестовая миграция)
их пребывания в остальное время года (места нагула) прежде всего благоприятными условиями для развития икры (подходящий субстрат, оптимальные температуры, достаточное содержание кислорода) и меньшим числом врагов. Обособление мест нагула и мест нереста расширяет использование видом среды, обеспечивая оптимальные условия для разных возрастных групп с их неодинаковыми потребностями. Некоторые морские виды (сельдь черноспинка и др.), видимо, раньше нерестившиеся, как и их родичи, в морях, постепенно перешли к нересту в реках, где меньше врагов, сохранив места нагула в море. Лососевые, некоторые карповые и осетровые рыбы, видимо, перво
начально были пресноводными, а затем, сохранив места нереста в реках, переменили места нагула на более кормные в морях. Речные угри, наоборот, для нереста уходят в море (см. рис. 100). Некоторые морские глубоководные рыбы для икрометания поднимаются в поверхностные слои, где вылупившаяся молодь лучше обеспечена кормом. Такие вертикальные нерестовые миграции совершают и байкальские голомянки.
 Центральная нервная система и органы чувств. У костных рыб относительные размеры головного мозга в большинстве случаев крупнее, чем у хрящевых рыб. В то же время основные функции разных отделов головного мозга в обоих классах, по существу, одинаковы. Передний мозг (telencephalon) костных рыб относительно невелик (рис. 129). Главную его массу составляют полосатые тела (corpora striata); нервные клетки в крыше переднего мозга отсутствуют или образуют лишь незначительные скопления (осетрообразные). Промежуточный мозг (diencephalon) довольно велик; на его спинной стороне образуется хорошо развитый эпифиз, а на брюшной стороне — гипофиз. Зрительные нервы образуют отчетливый перекрест (хиазму). Средний мозг (mesencephalon) у большинства костных рыб по размерам заметно превышает остальные отделы мозга и прикрывает промежуточный мозг. Сверху он подразделен на две зрительные доли, в которых оканчиваются волокна зрительного нерва; там проходит обработка зрительных восприятий. В брюшной части среднего мозга сосредота-
Центральная нервная система и органы чувств. У костных рыб относительные размеры головного мозга в большинстве случаев крупнее, чем у хрящевых рыб. В то же время основные функции разных отделов головного мозга в обоих классах, по существу, одинаковы. Передний мозг (telencephalon) костных рыб относительно невелик (рис. 129). Главную его массу составляют полосатые тела (corpora striata); нервные клетки в крыше переднего мозга отсутствуют или образуют лишь незначительные скопления (осетрообразные). Промежуточный мозг (diencephalon) довольно велик; на его спинной стороне образуется хорошо развитый эпифиз, а на брюшной стороне — гипофиз. Зрительные нервы образуют отчетливый перекрест (хиазму). Средний мозг (mesencephalon) у большинства костных рыб по размерам заметно превышает остальные отделы мозга и прикрывает промежуточный мозг. Сверху он подразделен на две зрительные доли, в которых оканчиваются волокна зрительного нерва; там проходит обработка зрительных восприятий. В брюшной части среднего мозга сосредота-

Рис. 129. Головной мозг форелн сверху и снизу (по Видерсгейму):
/ — обонятельная луковица, 2 — мозжечок, 3 — перекрест зрительных нервов, 4 — промежуточный мозг, 5 — эпифнз, 6 — гипофиз, 7 — зрительные долн среднего мозга, 8 — средний мозг, 9 — продолговатый мозг, 10 — спинной мозг» II — передний мозг; /—К — головные нервы
чиваются центры связи с мозжечком, продолговатым и спинным мозгом, в которых осуществляется суммация и сопоставление восприятий. У подвижных видов рыб мозжечок (cerebellum) хорошо развит, он частично налегает на средний мозг и почти полностью прикрывает продолговатый мозг. У малоподвижных рыб относительные размеры мозжечка уменьшаются. Осуществляя взаимодействие восходящих путей спинного и продолговатого мозга и нисходящих путей среднего мозга, мозжечок обеспечивает контроль над тонусом соматической мускулатуры, координацией движения и сохранением равновесия. Продолговатый мозг (myelencephalon seu medulla oblongata) по форме, относительным размерам и функции сходен с продолговатым мозгом хрящевых рыб.
У двоякодышащих, кистеперых и многоперообразных рыб пропорции отделов головного мозга отличаются от описанных выше и характерных для остальных костных рыб. Передний мозг у них крупнее остальных отделов; он разделяется на правое и левое полушария. В области желудочков нервные клетки подстилают крышу полушарий. Средний мозг и мозжечок имеют относительно небольшие размеры. Эти отличия, видимо, связаны с большей ролью обоняния, по сравнению со зрением, и малой подвижностью этих рыб.
Как и у хрящевых, от головного мозга костных рыб отходит 10 пар головных нервов (рис. 129 и 34; см. с. 66).
Спинной мозг (medulla spinalis) костных рыб по строению сходен со спинным мозгом хрящевых рыб. Он лежит в канале, образованном верхними дугами позвонков. Автономность функционирования спинного мозга у костных рыб выражена слабее, чем у хрящевых рыб.
Как и у хрящевых рыб, в ориентации и общении костных рыб особенно важную роль играют органы химического чувства (обоняние, вкус). Оба канала получения химической информации взаимодействуют, дополняя друг друга.
Обонятельные мешки костных рыб имеют внутри хорошо развитые складки обонятельного эпителия. Кожистый клапан разделяет ноздрю на два отверстия: у плывущей рыбы вода заходит в обонятельный мешок через переднее и выходит через заднее носовое отверстие У части рыб (плотва, угри и др.) ток воды через обонятельный мешок усиливается мерцанием ресничек, усеивающих его внутреннюю поверхность. Хотя обонятельные доли и весь передний мозг у большинства костных рыб относительно невелик (сильнее развиты у кистеперых, двоякодышащих и многоперообразных), пороги обонятельной чувствительности очень низки. Морские налимы —Gaidropsarus, сидевшие в бассейне емкостью 300 л, тотчас реагировали на прибавление 2,5 л воды из 12-литровой банки, в которой лишь 5 мин находилась одна рыбка атеринка. Еще ниже порог чувствительности у угря: запах воспринимается при наличии 15—50 тыс. молекул в 1 л (при-
1 У Двоякодышащих и части ископаемых кистеперых рыб от диа обонятельного мешка отходит каиал, открывающийся в ротовую полость внутренней иоздрей— хоаиой. Вода поступает в обонятельные мешки через наружные ноздри, а выходит через хоаиы.
мерно соответствует чутью собаки). Биологическое значение обоняния крстных рыб многообразно. Эксперименты показали, что рыбы хорошо распознают запахи не только чужих видов, но и различных особей своего вида. Это позволяет стайным рыбам не терять свою стаю, одиночным — находить партнера при размножении и т. п. Распознавание запаха чужого вида позволяет уклониться от нападения хищника или уловить приближение добычи. У многих рыб, особенно стайных, в особых бокаловидных клетках кожи содержится так называемое «вещество страха» или «опасности». При поранении кожи оно попадает в воду и воспринимается другими рыбами как сигнал опасности. Под действием такого химического сигнала стая уплотняется, что затрудняет нападение хищника, или, наоборот, рассеивается, и рыбы затаиваются. Велико значение обоняния и «химической памяти» у мигрирующих рыб. Так, тихоокеанские лососи выклевываются из икры в реках, проводят там несколько месяцев на стадии личинки и малька, а затем скатываются в море и кочуют. Через несколько лет, достигнув половой зрелости, рыбы мигрируют на нерест, проплывая сотни километров, и находят родные реки по запаху их вод *. Меченьем личинок было установлено, что из 13 тыс. выловленных на нерестилищах рыб 34% вошли точно в те реки и ручьи, где они выклюнулись из икры, 65 — в соседние и лишь 1 % рыб был пойман на значительном удалении от мест меченья.
При поисках и распознавании пищи важное значение имеет вкус. Вкусовые почки (скопление чувствующих клеток с оканчивающимися на них веточками головных нервов) расположены в слизистой оболочке ротовой области (щука, колюшки), у многих рыб они рассеяны и по всей поверхности тела, концентрируясь на голове, усиках и иногда на удлиненных лучах плавников. Экспериментально показано, что рыбы различают сладкое, соленое, кислое, горькое. Нитевидные грудные плавники некоторых придонных рыб высокочувствительны к механическим раздражениям и экстрактам из различных морских беспозвоночных. Острота вкуса связана с экологическими особенностями вида: пищевой специализацией^типом местообитаний, степенью развития других рецепторов. Слепая пещерная рыбка Anoptichtys распознает раствор глюкозы при концентрации 0,005%.
Как и у хрящевых, у костных рыб хорошо развиты органы боковой линии (сейсмосенсорные органы). У большинства костных рыб канал боковой линии проходит в толще кожи вдоль средней линии тела, а на голове сильно ветвится, образуя сложную сеть. Многочисленные мелкие отверстия, пронизывающие чешуи (см. рис. 41), сообщают полость канала с внешней средой. Расположенные на стенках канала скопления снабженных ресничками чувствующих клеток иннервируются боковой ветвью блуждающего нерва (X пара). Органы боковой линии воспринимают даже слабые перемещения частиц воды и инфра- звуковые колебания. С помощью этих органов рыбы ориентируются в потоках воды, воспринимают приближение или удаление добычи,
1 Последний определяется химическим составом воды, выделениями растений и постоянных обитателей реки.
хищника или партнера по стае, избегают столкновения с подводными предметами. Тонкость такой ориентации настолько велика, что ослепленные рыбы плавают с обычной скоростью, не натыкаясь на препятствия; они поддерживают строй в стае и успешно схватывают плавающую добычу.
К механорецепторам относятся и органы осязания. Они представлены расположенными в коже осязательными тельцами (скоплениями чувствующих клеток), рассеянными по всей поверхности тела и обычно более многочисленными на губах, усиках и лучах плавников. С их помощью рыбы воспринимают раздражения от прикосновения твердых тел. Температура воды воспринимается с помощью терморецепторов — рассеянных в поверхностных слоях кожи свободных нервных окончаний, видимо, различающихся по реакциям на разную температуру. Костные рыбы способны различать перепады температуры в 0,4 °С.
Рыбы способны улавливать изменения магнитного и электрического полей. При низком напряжении электрического поля они обычно ориентируют тело по силовым линиям, при увеличении напряжения двигаются к аноду; при возрастании напряжения наступает гальванонаркоз. Сильные электрические поля, возникающие у линий высокого напряжения, оказывают на поведение рыб угнетающее влияние. Электрические поля образуются вокруг тела любой рыбы, но они настолько слабы, что рецепторами не воспринимаются. Когда неэлектрические рыбы держатся группами (стаями), происходит суммация их индивидуальных электрических полей и образуется электрическое поле стаи, достаточное для восприятия отдельными членами. Оно играет важную роль в согласовании поведения отдельных членов стаи и их пространственной ориентации. Рецепторы электромагнитного чувства (рис. 130) расположены преимущественно на голове.

 />
/> 
 Среди костистых рыб есть виды, имеющие специальные электрические органы, у отдельных видов — большой мощности. В большинстве случаев, как и у скатов, они представлены участками преобразованной поперечнополосатой мускулатуры (с. 166). У рыб из отряда клюворылообразных, населяющих мутные водоемы тропической Аф-
Среди костистых рыб есть виды, имеющие специальные электрические органы, у отдельных видов — большой мощности. В большинстве случаев, как и у скатов, они представлены участками преобразованной поперечнополосатой мускулатуры (с. 166). У рыб из отряда клюворылообразных, населяющих мутные водоемы тропической Аф-
рики, небольшие электрические органы расположены на хвостовом стебле; их разряды создают вокруг рыбы электрическое поле, изменение которого позволяет рыбам оценивать характер предметов на расстоянии в несколько метров и тем самым обеспечивает ориентировку и поиск добычи в мутной воде (рис. 131). Электрические поляслабо электрических рыб служат средством разграничения кормовых участков, предотвращая вторжение на Рис. 131. Электрическое поле гимнар- занятую территорию. В спокойном ха (по Лиссман, 1963) состоянии мормир—Mormyrus кап
 ните генерирует 1 импульс в 1 с, а при беспокойстве — до 30 импульсов в 1 с. Плавающий гимнарх — Gymnarchus niloticus генерирует до 300 импульсов в 1 с напряжением в несколько вольт. Рецепторами служат кожные нервножелезистые клетки, расположенные преимущественно на спине и в каналах боковой линии.
ните генерирует 1 импульс в 1 с, а при беспокойстве — до 30 импульсов в 1 с. Плавающий гимнарх — Gymnarchus niloticus генерирует до 300 импульсов в 1 с напряжением в несколько вольт. Рецепторами служат кожные нервножелезистые клетки, расположенные преимущественно на спине и в каналах боковой линии.
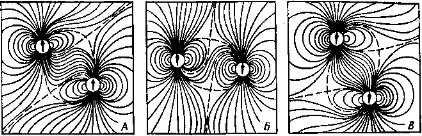
У части населяющих прибрежные тропические воды звездочетов Uranoscopidae (окунеобразные) на голове есть небольшие электрические органы. У звездочетов p. Astroscopus электрический орган, образованный преобразованными мышцами глаз, способен генерировать разряды напряжением до 50 В. Звездочет лежит на дне, полузакопавшись в грунт; когда мелкая рыбешка подплывает близко, следует разряд, и оглушенная добыча падает прямо в открывающийся громадный рот хищника. У электрических угрей (подотр. Gymnotoidei отряда карпообразных), населяющих пресные водоемы Южной Америки, расположенные по бокам тела электрические органы генерируют до 20—50 импульсов в 1 с и создают электрическое поле, которое обеспечивает ориентировку рыбы (глаза у нихЪчень малы). У электрического угря — Electrophorus electricus (рис. 132), достигающего 2 м длины, большие лентовидные электрические органы помимо низковольтных «ориентировочных» импульсов генерируют сильные разряды (с. 194). У живущего в водоемах Западной Африки электрического сома — Malapte- rurus electricus электрический орган в виде сплошного студенистого подкожного слоя окружает все туловище (рис. 132); он используется как орган ориентации, защиты и нападения (с. 196).
Зрение рыб — важное средство ближней ориентации и связи рыб в стаях.
Глаза костных рыб имеют уплощенную роговицу и шаровидный хрусталик (рис. 133). Подвижных век нет. Относительные размеры глаз в пределах класса варьируют. У живущих в условиях плохой освещенности (мутная вода, большие глубины и т. п.) видов глаза либо относительно очень крупные (у некоторых рыб приобретают «телескопическую» форму), либо, наоборот, очень малы, рудиментарны,4 а у некоторых пещерных рыб скрыты под кожей; в последнем случае
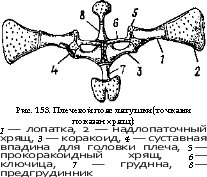
Рис. 132. Расположение электрических органов у разных рыб (по Хардеру, 1965):
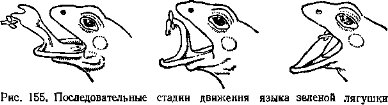 А — электрический скат — Torpedo; Б — обыкновенный скат — Raja;, В — электрический угорь — Electrophorus, Г — электрический сом — Malapterurus; Д — иож-рыба — Gymnotus; Е — гимнарх — Gymnarchus’, Ж — звездочет — Aslroscopus
А — электрический скат — Torpedo; Б — обыкновенный скат — Raja;, В — электрический угорь — Electrophorus, Г — электрический сом — Malapterurus; Д — иож-рыба — Gymnotus; Е — гимнарх — Gymnarchus’, Ж — звездочет — Aslroscopus
сильно развиты другие органы рецепции (боковая линия, обоняние, электрические органы и т. д.). В зависимости от образа жизни и пищевой специализации положение глаз изменчиво: у донных видов (см. рис. 98) они смещены к верхней стороне головы, у камбал оба глаза лежат на одной стороне головы (см. рис. 108). У выползающего на берег илистого прыгуна (см. рис. 98) крупные глаза как бы приподняты над головой и очень подвижны; у охотящейся на поверхности воды четырехглазки (см. рис. 102) крупные глаза с двойным зрачком и овальным хрусталиком позволяют одновременно видеть и в воздухе, и в воде. У большинства видов глаза размещены по бокам головы. Зрение преимущественно монокулярное: при поле зрения каждого глаза в 160—170° по горизонтали и около 150° по вертикали поле бинокулярного зрения составляет всего 20—30° (рис. 134).
В хрящевой склере иногда образуются мелкие окостенения. Наружная часть сосудистой оболочки обособляется в серебристую оболочку. У многих хрящевых и костных рыб на внутренней поверхности сосудистой оболочки образуется зеркальце или тапетум — слой плоских клеток, наполненных блестящими кристалликами гуанина; он отражает прошедшие через сетчатку световые лучи, что усиливает возможности зрения при слабой освещенности. Тонкий вырост сосудистой
Рис. 133. Схема вертикального разреза глаза костистой рыбы (по Паркеру):
/ — хрусталик, 2 — роговица, 3 — склера, 4 — сосудистая оболочка, 5 — серебристая оболочка (обособившийся наружный слой сосудистой оболочки), 6 — радужина, 7— пигментная оболочка, 8 — сетчатка, 9 — серповидный отросток, 10 —стекловидное тело, // — зрительный нерв
Рис. 134. Схема зрительных полей рыбы в горизонтальной плоскости (вид сверху) (по В. Р, Протасову):
I — поле зрения каждого глаза; II — поле монокулярного зрения; III — поле бинокулярного зрения; х — передняя и задняя слепые зоны
оболочки — серповидный отросток пронизывает сетчатку и стекловидное тело, прикрепляясь к хрусталику. При сокращении серповидного отростка хрусталик несколько сдвигается в глубь глаза; так осуществляется его аккомодация (настройка на резкость). Тонкий пигментный слой у видов с хорошо развитым тапетумом выражен слабо.
У большинства костных рыб сетчатка смешанная: содержит и палочки, и колбочки. При сильном свете увеличивающиеся' отростки пигментных клеток закрывают палочки, а колбочки подтягиваются к своим ядрам (установка на свет; рис. 135). При перемещении рыбы в темноту сокращаются пигментные клетки и прикрывают приблизившиеся к ним концевые членики колбочек; палочки подтягиваются к ядрам (установка на темноту;' рис. 135). Подобная ретиномоторная реакция (свойственная и всем остальным позвоночным) обеспечивает адаптацию зрения к меняющейся освещенности. Количество палочек и колбочек и их соотношение в сетчатке разных видов рыб сильно варьирует.
Рыбы воспринимают световые волны длиной в 400—750 нм. Максимумы световой чувствительности у пресноводных рыб по сравнению с морскими несколько сдвинуты в длинноволновую часть спектра. Большинство костных рыб имеют цветное зрение (некоторые, видимо, примерно в таких же пределах, как и человек). Глаза используются как рецепторы ближней ориентации (не далее 10—15 м); при равных условиях крупные рыбы имеют большую остроту зрения. Зрение играет важную роль при поисках пищи, уходе от опасности и при внутривидовом общении: при контактах в стае, поисках партнера для размножения, у некоторых рыб — при согласованном поведении молоди и родителей. Цветное зрение позволяет использовать особенности
окраски для распознавания особей своего вида и в некоторых случаях их физиологического состояния.
 Орган слуха и равновесия костных рыб, как и хрящевых, представлен только внутренним ухом, заключенным в хрящевую капсулу, наружные стенки которой окостеневают. Перепончатый лабиринт, или собственно внутреннее ухо, образовано лежащими во взаимоперпендикулярных плоскостях тремя хорошо развитыми полукружными каналами, отходящими от овального мешочка (вестибулярный аппарат или орган равновесия; см. рис. 39), лежащий ниже круглый мешочек имеет более или менее четко выраженный полый вырост — лагену — и служит собственно органом слуха. Отходящий от круглого мешочка тонкий эндолимфатический проток заканчивается слепо. В полости лагены, круглого и овального мешочков лежат отолиты, или слуховые камешки; они образованы кристалликами углекислой извести, скрепленными органическим веществом (по их слоистости, видной на шлифах, у многих видов можно определить возраст рыбы).
Орган слуха и равновесия костных рыб, как и хрящевых, представлен только внутренним ухом, заключенным в хрящевую капсулу, наружные стенки которой окостеневают. Перепончатый лабиринт, или собственно внутреннее ухо, образовано лежащими во взаимоперпендикулярных плоскостях тремя хорошо развитыми полукружными каналами, отходящими от овального мешочка (вестибулярный аппарат или орган равновесия; см. рис. 39), лежащий ниже круглый мешочек имеет более или менее четко выраженный полый вырост — лагену — и служит собственно органом слуха. Отходящий от круглого мешочка тонкий эндолимфатический проток заканчивается слепо. В полости лагены, круглого и овального мешочков лежат отолиты, или слуховые камешки; они образованы кристалликами углекислой извести, скрепленными органическим веществом (по их слоистости, видной на шлифах, у многих видов можно определить возраст рыбы).
 Принципы работы органа слуха и равновесия описаны выше (с. 73). У части костных рыб возникает связь плавательного пузыря с перепончатым лабиринтом: слепые отростки плавательного пузыря примыкают к затянутым перепонкой окошкам перилимфатической полости (некоторые окунеобразные, трескообразные) или образуется веберов аппарат — система косточек, соединяющих стенку плавательного пузыря с перилимфатической полостью внутреннего уха (карпообразные, сомообразные). Благодаря этому перепончатый лабиринт служит рецептором, улавливающим изменение давления в плавательном пузыре, а плавательный пузырь выполняет функцию резонатора и тем самым увеличивает остроту слуха.
Принципы работы органа слуха и равновесия описаны выше (с. 73). У части костных рыб возникает связь плавательного пузыря с перепончатым лабиринтом: слепые отростки плавательного пузыря примыкают к затянутым перепонкой окошкам перилимфатической полости (некоторые окунеобразные, трескообразные) или образуется веберов аппарат — система косточек, соединяющих стенку плавательного пузыря с перилимфатической полостью внутреннего уха (карпообразные, сомообразные). Благодаря этому перепончатый лабиринт служит рецептором, улавливающим изменение давления в плавательном пузыре, а плавательный пузырь выполняет функцию резонатора и тем самым увеличивает остроту слуха.
Костные рыбы воспринимают звуковые волны частотой от 16 до 12 ООО Гц. Рецептором служат чувствительные поля лагены и круглого мешочка, а возможно, и овального мешочка. Низкочастотные звуки не выше 500—600 Гц, видимо, могут восприниматься и органами боковой линии. Следует напомнить, что в воде звуки распространяются со значительно большей скоростью, чем в воздухе (около 1500 м/с против 330 м/с) и на большие расстояния. Поэтому звуковая ориентация для водных животных, в том числе и для рыб, очень важна. Звукопроводимость тканей тела рыб близка к звукопроводимости воды, и поэтому восприятие звуков возможно при относительно простом строении органа слуха.
В последние десятилетия выяснилось, что ходячее выражение «нем, как рыба» не соответствует действительности. Рыбы издают разнообразные звуки, воспринимаемые особями того же и других видов, как сигналы определенного значения. Механические или неспецифические звуки, возникающие при различных действиях рыбы (плавании, дыхании, еде), тоже имеют сигнальное значение. Звуки челюстей при схватывании добычи и при перетирании пищи привлекают других особей своего вида и хищников, звуки питания хищных рыб вызывают оборонительную реакцию у мирных рыб: бегство, затаивание, уплотнение стаи и т. п. Привлекает хищников н звук трепетания или «крики боли» схваченной рыбы. Более определенное видоспецифическое значение имеют звуки, специально производимые рыбой в определенной ситуации: при встрече партнеров в период размножения (нерестовые сигналы, включающие призыв особи другого пола, опознавание и стимуляцию выделения половых продуктов), сигналы предупреждения, и угрозы при защите отложенной икры или охране своей территории и т. п. Эти специфические сигналы по звучанию могут быть очень разнообразными. Скрипучие и скрежещущие звуки издаются при произвольном трении друг о друга свободных костных лучей плавников, костей челюстей и жаберной крышки, пояса грудных плавников и т. д.
Особенно разнообразны звуки, издаваемые с помощью плавательного пузыря: они напоминают барабанный бой, хлопки, свист, ворчание, кудахтанье, жужжание, стоны и т. д. У рыбы-жабы (батрахо- образные) на внешней стенке сердцевидного плавательного пузыря расположены плоские барабанные мышцы, а полость пузыря разделена плотной диафрагмой с отверстием посредине (рис. 136, А)\ издает звуки, похожие на хрюканье и пароходные гудки. У терапонов (окунеобразные) барабанные мышцы прикрепляются к затылочному отделу черепа и к стенкам передней части плавательного пузыря (рис. 136, Б), звуки напоминают хлопки или барабанную дробь. Низкие ворчащие звуки издает макрурус (трескообразные), у которого лентовидные барабанные мышцы лежат ' по бокам плавательного пузыря (рис. 136, В). Звуковые вибрации стенок плавательного пузыря могут вызывать колебания косточек веберова аппарата, костей пояса грудных плавников или ритмичные удары плавниковых лучей по телу. Пока не выяснены механизмы звучания у бычков, белуги и ряда других рыб. Частоты издаваемых рыбами звуков лежат в пределах 20—12 000 Гц, т. е. в пределах чувствительности их слухового аппарата. Наиболее разнообразные акустические сигналы свойственны сумеречным и придонным рыбам, рыбам со сложной популяционной структурой.
Таким образом, костные рыбы имеют разнообразные органы чувств. При ориентировке и при поисках пищи почти всегда используются многие рецепторы. Так, нарушение положения тела, например при ударе волны, воспринимается глазами, боковой линией, полукружными каналами и чувствительными полями круглого и овального мешочков (участками, к которым прилегают отолиты), плавательным пузырем и осязательными тельцами. При отыскании подвижной добычи используются зрение, органы боковой линии и электрические органы,
Рис. 136. Звуковые органы костистых рыб (по Протасову). А — рыба-жаба — Opsanus tau; Б — терапои— Therapoti\ В — макрурус — Macrurus berglax:
I — плавательный пузырь, 2 — барабаииая мышца. 3 — диафрагма о отверстнем, 4 — череп, 5 — первый позвонок, 6 — плавательный пузырь, 7 — барабанная мышца, 8 — плавательный пузырь» 9 — барабаииая мышца (внизу — поперечный разрез)
обоняние и осязание помогает находить неподвижную пищу, а захват и проглатывание пищи проходит под контролем зрения и вкусовых и осязательных телец, расположенных на усиках и в ротовой полости.
Поведение и образ жизни. Поведение и связанная с ним популяционная структура костных рыб довольно сложны. Богатство безусловнорефлекторной (врожденной) основы нервной деятельности рыб находит свое выражение в сложных инстинктах, обеспечивающих поиски и добывание пищи, размножение (нерестовые сигналы, выбор оптимальных мест для нереста, разнообразные формы заботы о потомстве), миграции, охрану индивидуальных участков или стайную организацию, обеспечивающую освоение больших территорий. Костные рыбы способны вырабатывать условные рефлексы на цвета, форму и размеры предметов, на звуки и другие раздражители. Все это определяет существование определенных и довольно различных у разных видов структур популяций. Сложные формы поведения рыб связаны не только с продолговатым и средним мозгом, но и с полосатыми телами переднего мозга. Так, цихлиды (окунеобразные) после удаления переднего мозга распознают особей другого пола, изучают новую территорию, но не могут откладывать икру и оплодотворять ее, теряют способность объединяться в стайки. У рыбок гемихромис способность охранять икру не восстановилась даже через 19 месяцев после повреждения полосатых тел.
Территориальное поведение сопровождается образованием различных внутрипо- пуляционных группировок. Их исходной формой, видимо, служит «выводок» — молодь, вылупившаяся из одной кладки. Только что выклюнувшиеся личинки не обращают внимания друг на друга, но уже через 2—3 дня обычно сближаются и повторяют движение соседей, составляя единую стайку с согласованным поведением (рис. 137). Этому способствует подражательное поведение, опирающееся на присущую молоди многих видов особую сигнальную «стайную» окраску — отчетливый рисунок или контрастные цветные пятна, — служащую опознавательными ориентирами. «Выводки» мальков обычно вскоре объединяются в большие стаи (элементарные популяции), состоящие из рыбок, развивавшихся совместно и обладающих сходным физиологическим состоянием и размерами. Такие группировки часто сохраняются до полового созревания. При движении в стае рыбы определенным образом подстраиваются друг к другу, обеспечивая гидродинамически благоприятное расположение (рис. 138). Упорядоченность размещения рыб в стае иногда сохраняется и во время отдыха.
 Преимущества стайной жизни так называемых «мирных» рыб несомненны: стая быстрее находит скопления пищи, легче обнаруживает приближение врага; при появлении последнего рыбы сбиваются в столь плотную группу, что даже крупному хищнику трудно оторвать жертву, или, наоборот, рассыпаются, дезориентируя хищника. Хищники-рейдеры образуют «рассеянные стаи», в которых одиночки или группы из нескольких рыб держатся разобщенно, но в пределах «видимости» (зрительной или акустической). Такое построение облегчает поиск подвижной добычи, ее атаку и захват. Многие виды рыб живут стаями в течение всей жизни (сельдевые, тресковые, карпообразные, окунеобразные и др.). Другие объединяются при кормовых и нерестовых миграциях, но распадаются на мелкие группировки на кормовых пастбищах и на нерестилищах, где нередко самки занимают отдельные участки для икрометания, иногда охраняемые и после откладки икры либо самкой (тихоокеанские лососи), либо самцом (некоторые тропические сомы и др.). Такие пресноводные хищники, как сом и щука, и ряд донных морских рыб (удильщикообразные, мурены из угреобразных, бычки-подкаменщики из скорпенообразных и др.) ведут одиночный образ жизни.
Преимущества стайной жизни так называемых «мирных» рыб несомненны: стая быстрее находит скопления пищи, легче обнаруживает приближение врага; при появлении последнего рыбы сбиваются в столь плотную группу, что даже крупному хищнику трудно оторвать жертву, или, наоборот, рассыпаются, дезориентируя хищника. Хищники-рейдеры образуют «рассеянные стаи», в которых одиночки или группы из нескольких рыб держатся разобщенно, но в пределах «видимости» (зрительной или акустической). Такое построение облегчает поиск подвижной добычи, ее атаку и захват. Многие виды рыб живут стаями в течение всей жизни (сельдевые, тресковые, карпообразные, окунеобразные и др.). Другие объединяются при кормовых и нерестовых миграциях, но распадаются на мелкие группировки на кормовых пастбищах и на нерестилищах, где нередко самки занимают отдельные участки для икрометания, иногда охраняемые и после откладки икры либо самкой (тихоокеанские лососи), либо самцом (некоторые тропические сомы и др.). Такие пресноводные хищники, как сом и щука, и ряд донных морских рыб (удильщикообразные, мурены из угреобразных, бычки-подкаменщики из скорпенообразных и др.) ведут одиночный образ жизни.
При недостатке пищи в водоеме нередко возникает каннибализм — поедание своей икры и молоди. В некоторых случаях он может стано-
Рис. 138. Силовое поле вокруг двигающихся рыб. А — рыбы взаимно притягиваются; Б — рыбы не влияют друг на друга; В — рыбы взаимно отталкиваются (по Шулей-
кину)
виться даже нормой. В отдельных озерах Западной Сибири крупные особи обыкновенного окуня питаются преимущественно мелкими окуньками, которые в свою очередь живут за счет планктона — пищи, не доступной для крупных окуней. Это дает возможность популяции существовать в водоемах, где для взрослых особей нет другого корма.
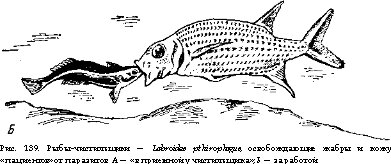
Особо сложные внутривидовые и межвидовые отношения характерны для рыб, населяющих тропические водоемы и коралловые рифы. Им сопутствует огромное разнообразие цветов и рисунков в окраске, развитие звуковой сигнализации и демонстрационного поведения, сложные формы брачного поведения и причудливый гермафродитизм, забота о потомстве (постройка гнезд, охрана икры и молоди). Такая внутривидовая (популяционная) организация упорядочивает использование пространства с его жизненными ресурсами. Примером усложнения взаимоотношений в богатых видами биоценозах тропических морей может служить существование мелких «рыб-чистилыциков», собирающих паразитов с кожи, поверхности глотки и жабр более крупных рыб — «пациентов». Последние иногда собираются к местам обитания «чистильщиков» большими группами, карикатурно напоминающими пациентов, терпеливо ожидающих в приемной «врача» (рис. 139). Сами чистильщики обычно ярко окрашены, что предупреждает нападение на них. Одна из таких чистильщиков — рыбка Labroi- des dimidiatus, обитающая в тропиках Тихого океана, — представляет быстро и обратимо реверсирующего гермафродита, способного сменить состояние самца на состояние самки или обратно в течение нескольких минут. Они живут группами, состоящими из главы — крупного самца и гарема самок, среди которых крупные особи доминируют над мелкими. Самец активным поведением препятствует превращению самок в самцов, но при его гибели доминирующая самка немедленно превращается в самца, занимая его место (Робертсон, 1972). Отношения чистильщиков и обслуживаемых ими крупных рыб повели к появлению своеобразной мимикрии, когда мелкие хищники, по форме и окраске напоминающие чистильщиков, проникают в жаберные полости обманутых «пациентов», вырывая куски жабр.
Для видов с одиночным образом жизни часто характерны защитная окргска и мощное вооружение (шипы, острые лучи плавников, иногда
имеющие у основания ядовитые железы, и т. п.), у них обычно есть специальные звуковые сигналы, связанные с призывом самки и охраной территории.

 Особенности образа жизни и поведения рыб, принадлежащих к различным отрядам, описаны выше. Независимо от экологического облика или систематической принадлежности жизнь рыбы всегда состоит нз чередования в определенной последовательности событий, в своей совокупности образующих биологический цикл, тесно связанный с сезонными изменениями условий существования. Он состоит из размножения, нагула, подготовки к зимнему периоду и зимовки, после которой наступает сезон нового размножения. Отдельные этапы жизненного (сезонного) цикла рыб освещены при описании питания, размножения и других биологических явлений.
Особенности образа жизни и поведения рыб, принадлежащих к различным отрядам, описаны выше. Независимо от экологического облика или систематической принадлежности жизнь рыбы всегда состоит нз чередования в определенной последовательности событий, в своей совокупности образующих биологический цикл, тесно связанный с сезонными изменениями условий существования. Он состоит из размножения, нагула, подготовки к зимнему периоду и зимовки, после которой наступает сезон нового размножения. Отдельные этапы жизненного (сезонного) цикла рыб освещены при описании питания, размножения и других биологических явлений.
Важным элементом годового жизненного цикла многих видов рыб являются миграции — перемещения со сменой мест обитания. Миграции могут быть активными и пассивными. В первом случае рыбы активно движутся в избранном направлении, иногда преодолевая сильные течения и даже пороги (например, лососевые). При пассивных миграциях используется сила течения. Обе формы миграции обычно сочетаются: активная — у взрослых, пассивная — у личинок и молоди (миграции личинок сельди, угря и др.). Пассивная миграция имеет место и у некоторых малоподвижных пелагических рыб, живущих в районах круговых теплых течений (луна-рыба и др.).
Во время активных миграций рыбы ориентируются с помощью всех органов чувств. Считают, что особенно важны химические и температурные восприятия. Хотя рыбы способны различать даже небольшие, в доли градуса, перепады температур, основным направляющим, видимо, служит химическое чувство. Взрослые угри плывут в направлении повышающейся, а лососевые — понижающейся солености. Как указывалось выше, при входе в реку рыбы ориентируются с помощью химической памяти, сохранившей с личиночного периода запах «родной» реки. Наблюдения за поведением мигрирующих в море стай тихоокеанских лососей позволили предположить, что выбор общего направления к родному берегу осуществляется с помощью солнечной ориентации или астронавигации (предположение нуждается в проверке).



 В жизни многих рыб миграции разного типа сменяют друг друга (рис. 140). После нереста, совершая кормовые миграции, рыбы переходят на богатые кормом «пастбища». Часть видов держится на этих кормовых участках более или менее оседло до следующего цикла размножения, другие постоянно перемещаются в поисках пищи. С наступлением сроков очередного размножения начинаются нерестовые миграции. У многих пресноводных и некоторых морских рыб северных и умеренных широт после нагула происходят зимовочные миграции. На местах зимовок в состоянии минимальной активности рыбы переживают зимний период. В пресных водоемах местами зимовок обычно служат наиболее глубокие участки — «ямы». В нижнем течении Урала, Волги и других каспийских рек в «ямах» зимуют осетровые, вобла, лещ, сазан и судак. Рыбы «стоят» неподвижно, близко друг к другу, иногда в несколько слоев. Нагуливающаяся и нерестящаяся в Азовском море хамса осенью уходит на зимовку в Черное море. Дальневосточные камбалы в районе залива Петра Великого нагуливаются в прибрежных районах моря, а на зимовках концентрируются в немногих участках на глубинах около 100—150 м; они выбирают участки с положительной температурой и закапываются в ил, иногда в несколько слоев.
В жизни многих рыб миграции разного типа сменяют друг друга (рис. 140). После нереста, совершая кормовые миграции, рыбы переходят на богатые кормом «пастбища». Часть видов держится на этих кормовых участках более или менее оседло до следующего цикла размножения, другие постоянно перемещаются в поисках пищи. С наступлением сроков очередного размножения начинаются нерестовые миграции. У многих пресноводных и некоторых морских рыб северных и умеренных широт после нагула происходят зимовочные миграции. На местах зимовок в состоянии минимальной активности рыбы переживают зимний период. В пресных водоемах местами зимовок обычно служат наиболее глубокие участки — «ямы». В нижнем течении Урала, Волги и других каспийских рек в «ямах» зимуют осетровые, вобла, лещ, сазан и судак. Рыбы «стоят» неподвижно, близко друг к другу, иногда в несколько слоев. Нагуливающаяся и нерестящаяся в Азовском море хамса осенью уходит на зимовку в Черное море. Дальневосточные камбалы в районе залива Петра Великого нагуливаются в прибрежных районах моря, а на зимовках концентрируются в немногих участках на глубинах около 100—150 м; они выбирают участки с положительной температурой и закапываются в ил, иногда в несколько слоев.
Начало зимовочных миграций определяется физиологическим состоянием рыб (количеством накопленного жира, появлением «холодных» ферментов и др.) и изменением внешних условий (температуры, солености, содержания кислорода). При отсутствии жирового запаса зимовочная миграция может не состояться. Так, хамса, имеющая жирность менее 14%, не мигрирует из Азовского моря даже при резких понижениях температуры воды, при жирности 14—17% начинает миграцию при перепаде температур в 9—14 °С, а при жирности около 22% дружно мигрирует даже при небольшом снижении температуры. Иногда на зимовку уходят половозрелые популяции, а неполовозрелые особи продолжают кормиться всю зиму (лещ, судак и др.).
Динамика численности рыб отражает взаимодействие их популяций со средой. Ее характер обусловливается многими факторами: продолжительностью жизни особей данного вида, характером и темпом их размножения, обеспеченностью пищей, размерами смертности и изменчивостью факторов среды (колебаниями уровня воды, температуры и содержания кислорода, изменениями мощности и направления морских течений и др.). У моноциклических видов, размножающихся только раз, в конце жизни, численность обычно менее устойчива и подвержена большим колебаниям, чем у долго живущих полицикли- ческих видов. Устойчивость численности зависит от обеспеченности пищей; существенное влияние оказывают враги и пищевые конкуренты. Вследствие большей стабильности физико-химических условий в водной среде, особенно в морях, по сравнению с сушей колебания численности рыб происходят с меньшей амплитудой, чем у многих наземных животных. Но промерзание нерестилищ в суровые зимы катастрофически сказывается на отдельных популяциях дальневосточных лососей, а перетирание плавающими льдами икры — на урожайности сельдей Охотского моря.
В последнее время усиливающееся воздействие на динамику численности рыб оказывает хозяйственная деятельность человека: увеличивается промысел, возрастает загрязнение водоемов промышленными и другими отходами, строительство гидростанций меняет гидрологический режим и т. п.
Частота и амплитуда колебаний числа особей в популяциях изменяется в широких пределах. Так, уловы тихоокеанского палтуса — Н. hippoglossus за период 1925—1947 гг. колебались в двукратном размере, а уловы тихоокеанской сардины — Sardinops sagax в 1937— 1957 гг. изменялись от 2 млн. центнеров до нуля. Высокая численность идущей в реки тихоокеанского побережья горбуши обычно бывает через год, а у атлантического лосося — раз в 10—11 лет.
Промысел, изреживая популяцию, приводит к «омоложению» ее возрастного состава: уменьшается доля медленно растущих старших возрастных групп, а улучшение питания приводит к ускоренному росту и более раннему созреванию молодых рыб (рис. 141). Таким образом, промысел в определенном размере оказывает положительное влияние на воспроизводство популяции, усиливая интенсивность размножения и темп роста рыб. Однако если вылов превышает норму (различную для разных видов и районов), он уменьшает стадо произво
дителей и численность популяции начинает снижаться. Следовательно, для каждого вида и для данного водоема важно определить оптимальные нормы отлова, обеспечивающие постоянно высокие уловы за счет поддержания повышенной воспроизводительной способности популяции.
alt="" />Роль рыб в водных биоценозах.
Рыбы завершают многие цепи питания. Поэтому их роль в регуляции потоков энергии и круговороте веществ исключительно велика.
Воды занимают^около 72% поверхности земиого шара. В определенные периоды года в них развивается громадное количество фитопланктона, за счет которого живет зоопланктон. Оба они служат пищей планктоноядных рыб и молоди почти всех других рыб. Много рыб из разных групп питается донными беспозвоночными: червями, моллюсками, ракообразными, иглокожими и др. Крупные рыбы поедают более мелких рыб. В свою очередь рыбы или их молодь служат пищей для таких беспозвоночных, как медузы, паразитические черви, головоногие моллюски, крупные ракообразные, иглокожие, водные насекомые. Трупы рыб разлагаются бактериями, завершающими круговорот веществ.
Рыбы используются в пищу представителями всех классов позвоночных животных. Икру и особенно молодь поедают земноводные. Преимущественно рыбами питаются водные пресмыкающиеся: некоторые ужи, морские змеи, крокодилы. Среди птиц в той или иной степени питаются рыбами гагары, поганки, веслоногие, некоторые утки (особенно крохали), из хищных — скопа и орлан-белохвост, совы (рыбный филин), зимородки и др. Некоторые виды рыбоядных птиц гнездятся громадными колониями — птичьими базарами, — приуроченными к районам, богатым рыбой. Колонии на островах у берегов Перу образованы миллионами веслоногих птиц (бакланы, олуши), поедающих ежегодно сотни тысяч тонн мелкой рыбы, преимущественно перуанского анчоуса. Птичьи базары Севера (на берегах Исландии, Гренландии, Мурмана, Новой Земли, Чукотки, Командорских и Курильских о-вов) насчитывают иногда десятки и даже сотни тысяч гнездящихся птиц: чаек, чистиковых, в некоторых местах — бакланов и др. Помет массы птиц, попадающий в воду, обогащает ее азотистыми веществами, что способствует бурному развитию фитопланктона, на нем кормится зоопланктон, привлекающий рыб. Рыбы поедаются птицами... Такие крупные колонии обычно приурочены к районам столкновений холодных и теплых течений, создающим интенсивную вертикальную циркуляцию
водных масс и способствующих обогащению их минеральными веществами и кислородом, что обеспечивает интенсивное развитие фитопланктона. Довольно много потребителей рыбы и среди млекопитающих. Мелкими стайными рыбами питаются зубатые и некоторые усатые киты, разнообразные виды ластоногих. В пресных водоемах рыбу поедает выдра, норка, водяная землеройка-кутора. Во время массового хода рыбы ее ловят медведи, волки и другие «сухопутные» хищники.
Еще по теме Кровеносная система и кровообращение:
- Кровеносная система и кровообращение
- 14.4. КРОВЕНОСНАЯ СИСТЕМА
- КРОВЕНОСНАЯ СИСТЕМА
- Кровеносная система
- Кровеносная система
- Кровеносная система
- Кровеносная и выделительная системы
- Кровеносная система глаза.
- ГЛАВА 4. КРОВЕНОСНАЯ И ВЫДЕЛИТЕЛЬНАЯ СИСТЕМЫ
- Физиология кровообращения
- ПРЕПАРАТЫ, РЕГУЛИРУЮЩИЕ РИТМИКУ СЕРДЦА И КОРОНАРНОЕ КРОВООБРАЩЕНИЕ
- Малый круг кровообращения
- Артерии большего круга кровообращения
- ДВА АНАТОМИЧЕСКИХ ТРАКТАТА О КРОВООБРАЩЕНИИ
- ВТОРОЕ ПОСЛАНИЕ ИОАННУ РИОЛАНУ В котором опровергаются многие возражения против кровообращения
- 20.1.1.2. Сосальщики с одним промежуточным хозяином, обитающие в кровеносных сосудах
- ПЕРВОЕ ПОСЛАНИЕ ИОАННУ РИОЛАНУ Анатомическое исследование о кровообращении
- Глава 8 ПОНЯТИЯ О СИСТЕМЕ УДОБРЕНИЙ И ОСНОВНЫЕ ПОЛОЖЕНИЯ СИСТЕМЫ УДОБРЕНИЯ В СЕВООБОРОТЕ
- ПРИМЕНЕНИЕ БЕСПОДСТИЛОЧНОГО НАВОЗА КАК СОСТАВНАЯ ЧАСТЬ СИСТЕМЫ УДОБРЕНИЯ Основы системы удобрения
- Ценотическая система Ценотическая система — что это такое?