Взаимодействие морских бореальных биот Атлантики и Пацифики
Российским морским биологам наиболее известен контакт аркто-атлантической и тихоокеанской умеренных биот. На протяжении длительного периода Северный Ледовитый океан был связан с Атлантикой сначала через Тургайский пролив, а затем, после соединения Европы с Азией и отделения Европы от Америки — через пролив между Гренландией и Европой.
Шельфы Северной Атлантики и Северного Ледовитого океана, с одной стороны, и Северной Пацифики, с другой, располагались в примерно одних климатических условиях, площадь их была одного порядка, поэтому логично предположить, что в этих двух акваториях сформировался сходный по экологическим амплитудам набор ЦС.Поскольку атланто-арктическая и северо-тихоокеанская биоты произошли от разных биот, они были образованы каждая своими эндемичными таксонами, несмотря на сходные абиотические условия.
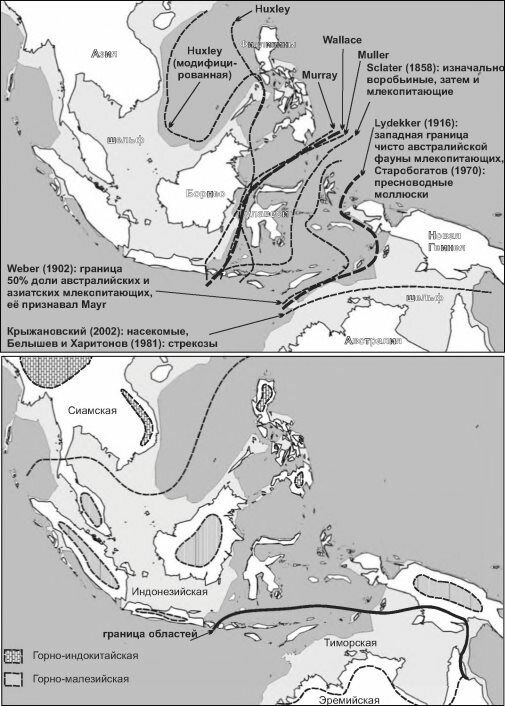
Рис. 4.43. Положение границы между австралийской и азиатской биотой по разным авторам. Верхняя карта — по разным группам животных (авторы указаны на карте), нижняя — по сосудистым растениям (скомпилировано мной по оригинальным картам из архива С.М. Разумовского), подписаны названия провинций, их границы даны пунктиром; Тиморская и Эремийская провинции принадлежат австралийским областям, остальные — Малайской области; горные и эремийская — провинции субтропических лесов, остальные — тропических. Горные провинции показаны схематично.
Берингов пролив открылся 5,32 млн. лет назад (Gladenkov et. al., 2002), впервые за 60 млн. лет, и аркто-атлантическая и северо-тихоокеанская биоты вступили в контакт. Тогда пролив был узким и мелководным, возможно, широкого обмена биотами в то время и не произошло. Вскоре пролив закрылся и вторично открылся примерно 3,5 млн.
лет назад. Условия в Северном Ледовитом океане тогда были сходны с существующими сейчас в Северной Атлантике, т. е. такими же, как в Северной Пацифике. Тем не менее, в результате контакта не образовалась единая северная биота (как произошло бы при правоте контину- алистов), напротив, каждая из биот сохранила свою самостоятельность.Всё же некоторый обмен видами между двумя биотами произошёл, хотя и неравноценный. Практически среди всех таксонов крупного ранга, по которым имеются данные, преобладала миграция из Тихого океана. Так, по данным G.J. Vermeji (1991), из 295 видов моллюсков, участвовавших обмене или непосредственно произошедших от таких видов, 261 вид имели тихоокеанское происхождение, аркто-атлантическое — только 34. Доля тихоокеанских видов в Атлантической биоте явно завышена, поскольку к видам, мигрировавшим в Атлантику из Пацифики через Арктику (или произошедшим от таких видов) G.J. Vermeji отнёс и большое количество видов, в действительности не выходящих за пределы Арктики. Всё же тенденция им подмечена верно. Другие группы дают сходные результаты: миграция преимущественно со стороны Пацифики наблюдалась среди десятиногих ракообразных (Spirontocaris и Crangon — рода безусловно тихоокеанского происхождения в Северной Пацифике 71 вид Spirontocaris и 14 видов Crangon, в Северной Атлантике соответственно — 4 и 2, общих обоим регионам — 9 и 1) и иглокожих (Дьяконов, 1945). Из рыб морские окуни несомненно вселились в Атлантику из Пацифики, тресковые же — наоборот, из Атлантики в Пацифику. Из водных млекопитающих тихоокеанское происхождение имеют отряд сирен, ушастые морские тюлени, моржи и морские выдры, но настоящие тюлени атлантического происхождения. Среди красных водорослей (Пере- стенко, 1994) из 9 арктическо-бореальных родов 8 — тихоокеанского происхождения и лишь один — атлантического, морские травы (Zostera и Phyllospadix) тихоокеанского происхождения.
Наиболее вероятной основной причиной этой асимметрии является различие в видовом богатстве фаун, участвовавших в обмене.
И в настоящее время (т. е. после совершившегося обмена) тихоокеанская шельфовая фауна вдвое богаче атлантической, весьма вероятно, что это различие было ещё больше до начала обмена. Часто видовое богатство связывают с площадью, на которой она существует (т. е. с потоком энергии через её ареал) или с временем существования биоты. Однако в данном случае эти объяснения, по-видимому, не подходят. Северо-пацифическая и аркто-атлантическая биоты имеют скорее всего одинаковый возраст. В конце палеоцена (57 млн. лет назад) произошло быстрое глобальное потепление. Поверхностная температура воды в Антарктике достигала 18°С. Это привело к исчезновению всех умеренно-теплолюбивых морских биот. Вновь условия для их возникновения возникли 25 млн. лет назад, с установлением существенного температурного градиента между экватором и полюсом, поэтому северо-пацифическая и североатлантическая биоты не могут быть древнее 25 млн. лет. Площади, занимаемые этими биотами, как указывалось выше, сходны. Логично предположить, что различия в видовом богатстве северо-атлантической и северо-тихоокеанской биот в значительной степени обусловлены различиями в видовым богатстве донорских биот. Атланто-арктическая фауна с юга она граничила с центрально-американской и восточно-атлантической тепловодными фаунами, тогда как северо-тихоокеанская — с той же центрально-американской (Панамского перешейка тогда не было) и индо-малайской. Восточно-атлантическая биота уже тогда была гораздо беднее индо-малайской.Другая причина — миграции нотальных биот. Поскольку охлаждение умеренных широт в Южном полушарии произошло, как минимум, на 10 млн. лет раньше, чем в Северном, нотальные биоты сформировались гораздо раньше. При наличии подходящих условий (а они были особенно благоприятны в периоды оледенений), происходил обмен бореальных и нотальных биот. Выше в этой главе (раздел «Причины и скорость образования
—
*?
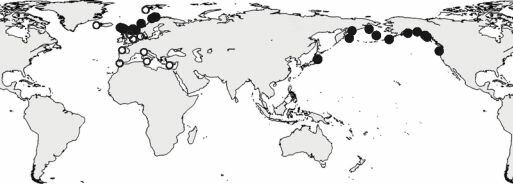
Рис.4.44.
Ареал полихеты Pectinaria auricoma.Чёрные кружки — наши данные, светлые — распространение по сводке Т. Holthe (1986).
новых видов и экосистем») было показано, что наиболее подходящий путь такого обмена — вдоль тихоокеанских берегов Америки, дополнительные аргументы приведены также в разделе о биполярных ареалах (глава 6).
Теоретически существует два возможных пути миграции тихоокеанской шельфовой биоты в Атлантику: вдоль Евразии и вдоль Северной Америки. Последний путь, начиная с работ S. Ekman (1935), Е.Ф. Гурьянова (1957) и, особенно, после работы К.Н. Несиса (1961), часто считают главным или даже единственным. Эту точку зрения подкрепляют двумя основными аргументами: (1) путь в Атлантику через Канадский арктический архипелаг короче, чем вдоль Евразии, к тому же, поверхностные течения в период миграции бореальных видов, скорее всего были теми же, что и ныне, т. е. шли из Атлантики вдоль Евразии, а от Чукотского моря вдоль Америки, Е.Ф. Гурьянова даже предполагает, что хребет Ломоносова выступал над поверхностью моря, делая невозможным евразийскуий миграционный путь; (2) существуют виды, обитающие в Пацифике и только в западной Атлантике, например краб Chinoecetes opilio и плоский ёж Echinarachnius рагта. Относительно первого аргумента можно отметить, что некоторые виды, проникшие в Атлантику из Пацифики не имеют пелагических личинок, поэтому направление течений для них несущественно. Относительно второго аргумента сам К.Н. Несис пишет, что в конце плиоцена «нынешние тихоокеанские-западноатлантические виды моллюсков обитали в Англии и Исландии» (1961: 895). Также имеются виды с тихоокеанско-восточноатлантическим ареалом, причём не только мелководные, как утверждает К.Н. Несис, но и преимущественно батиальные (например, полихета Pectinaria auricoma, рис. 4.44). Л.П. Перестенко (1994), исследовав современное распространение красных водорослей (т. е. видов верхнесублиторальных, на которых постулируемые К.Н. Несисом факторы должны были сказываться в наибольшей степени), пришла к прямо противоположному выводу: о преимущественно евразийском пути обмена.
Среди видов исследованной ею флоры нет видов, обитающих только в западной Атлантике и западной Пацифике, хотя есть виды, обитающие только в западной Пацифике и восточной Атлантике. Л.П. Перестенко указывает, что вселение западнотихоокеанских видов происходит в европейские, а не американские воды.G.J. Vermeji (1991) объясняет тихоокеанско-западноатлантические ареалы относительной бедностью западноатлантической биоты по сравнению с восточноатлантической. Это привело к появлению в западной Атлантике сравнительно бульших свободных ресурсов, которые и стали потреблять вселившиеся из Тихого океана виды. Поэтому, вероятно, более правильно другое объяснение: тихоокеанских видов в западной Атлантике больше не потому, что им было туда легче попасть, а потому, что им там было легче закрепиться. В частности, оказалось, что краб-стригун Chionoecets opilio способен пройти пешком из

Рис.4.45. Ареал краба Chionoecetes opilio.
Звёздочкой отмечено место поимки экспедицией на «Polarstem». По: В.А. Павлову (2000); В.В. Петряшову и др. (1993); Jadamec et al., 1999; Squires, 1968 и лич. сообщ. А.Н. Карасева
«С уверенностью нельзя назвать ни одного рода, выселившегося в Атлантику через север Канады» (Перестенко, 1994: 233).
Чукотского моря в Баренцево: в 1993 г. он был пойман на полпути (в море Лаптевых) на расстоянии в тысячи км от границы ближайшей части его современного ареала (рис. 4.45). Совершенно очевидно, что этот путь взрослая особь краба прошла пешком, причём по евразийскому пути. Каких-либо палеонтологических следов таких миграций, естественно, остаться не могло. Что касается западно- и восточноарктических ареалов, то ниже, при рассмотрении биогеографии Северного Ледовитого океана (глава 6), показано, что таких видов, по-видимому, в реальности нет.
Контакт тихоокеанской и атлантической шельфовых биот на юге Южной Америки
Шельф юга Южной Америки, на котором и ныне выходят ледники, в плейстоцене, несомненно, был занят ледником, лежавшим на дне, подобно тому, как это сейчас наблюдается в Антарктиде.
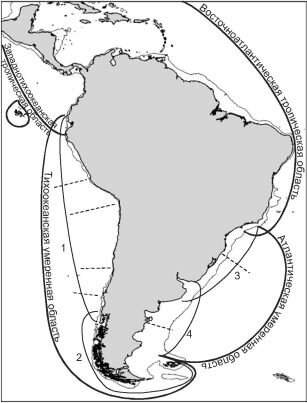
Этот лед-Рис. 4.46. Районирование южноамериканского шельфа по степени значимости зоогеографических границ. По В.Н. Семёнову (1982). Подобласти: 1 — Перуано-Чилийская; 2 — Южночилийская; 3 — Южнобра- зильско-Уругвайская; 4 — Аргентинская. Пунктиром показаны границы провинций, тонкая линия — изобата 200 м.
Районирование вдоль тихоокеанского берега подтверждено данными по распространению 504 видов полихет (Hernandez et. al., 2005), причём работы В.Н. Семёнова (1982) в списке литературы к статье нет. В.Н. Семёнов (1982) использовал данные по распространению 333 вида полихет, его данные были гораздо менее полными.
ник неизбежно стал барьером, разделившим единую ранее умеренную южноамериканскую шельфовую биоту на тихоокеанскую и атлантическую, по крайней мере те её компоненты, которые не были способны обходить этот барьер по материковому склону. Это разделение привело к формированию двух комплексов умеренных ЦС, подобно тому, как это было в разобранном выше примере флоры Северной Америки. Разрушение ледника устранило разделявший эти биоты барьер. Поскольку вступившие в контакт наиболее южные атлантическая и тихоокеанская шельфовые биоты сформировались в весьма сходных условиях, по континуалистской парадигме следовало бы ожидать широкого их обмена, результатом которого было бы образование единой южной умеренной биоты. Вероятно поэтому все авторы, занимавшиеся биогеографией этого района (Ekmam, 1935, 1953; Гурьянова, 1964; Briggs, 1995 и многие другие, обзор см. Семёнов, 1982) на своих схемах показывают единую южноамериканскую умеренную (нотальную) область. В.Н. Семёнов (1982), детально изучив распространение свыше полутора тысяч видов южноамериканского бентоса, показал, что такое представление весьма далеко от реальности. В действительности, несмотря на то, что ледниковый барьер исчез, атлантическая и тихоокеанская биоты по-прежнему весьма различны, что заставляет рассматривать их ареалы как отдельные области (рис. 4.46). Этот необъяснимый и неожидаемый с позиций континуализма эффект вполне тривиален с позиций структурализма.
Еще по теме Взаимодействие морских бореальных биот Атлантики и Пацифики:
- Взаимодействие неморских биот Северной и Южной Америк
- Аркто-бореальное
- Метод выделения конкретных биот
- Метод выделения типов биот
- Примеры контакта различных биот
- МЕЖОРГАНИЗМЕННЫЕ ВЗАИМОДЕЙСТВИЯ
- 3.6.5.2. Взаимодействия между генами в генотипе
- Морские перья
- Морские звезды
- Взаимодействие токсикантов с липидами.
- Взаимодействие токсикантов с нуклеиновыми кислотами.
- 6.3.1.3. Наследование признаков, обусловленных взаимодействием неаллельных генов
- Взаимодействия насекомого-фитофага и растения
- Модели межклеточного взаимодействия и формообразования
- РОД КАЛИФОРНИЙСКИЕ МОРСКИЕ ЛЬВЫ GENUS ZALOPHUS