Закон Харли - Вайнберга
В идеальной популяции, когда между всеми ее особями идет свободное скрещивание и нет заноса генетического материала из других популяций, неизбежно установится равновесие доминантных и рецессивных аллелей.
Допустим, что некоторый ген представлен двумя аллелями: доминантным А и рецессивным а, тогда частота встречаемости А может быть принята за q и частота аллеля а за р. Сумма этих частот равна единице: q + р = 1.При этих соотношениях частота аллелей в потомстве первого поколения будет следующей:
(рА 4 qa) • (рА + qa) — ppAA + pqAa + qpaA + qqaa.
После преобразований получаем 2 (pA + qa) = pZAA + 2pqAa + q2aa.
В дальнейших поколениях частота аллелей останется такой же. Это явление получило название закона Харди - Вайнберга, который гласит, что при равноценности всех генов первоначальное соотношение аллелей независимо от их исходной частоты сохраняется во всех последующих поколениях. Он базируется на закономерностях, открытых Менделем, и справедлив лишь для популяций, находящихся в стабильных условиях.
Популяция, в которой соотношение генотипов соответствует уравнению Харди - Вейнберга, называется генетически равновесной. Равновесие частот генотипов в популяции поддерживается из поколения в поколение, если не нарушаются условия выполнения статистических закономерностей, го есть если скрещивания случайны, жизнеспособность особей с разными генотипами одинакова, не происходят изменения частот аллелей за счет мутаций, миграций или каких-либо других факторов. Разумеется, генетическая равновесность имеет среднестатистический харе ктер, у отдельных особей или групп особей наблюдаются отклонения от среднего типа.
В реальных популяциях отклонения от равновесия, а следовательно, и от закона Харди - Вайнберга могут быть весьма существенными. Известно несколько факторов, которые могут нарушать равновесие генотипов в популяции.
Это мутационный процесс, низкая численность особей, избирательный характер скрещиваний, занос особей из других популяций и естественный отбор. В результате соотношение между встречаемостью гомо- и гетерозиготных особей изменится, и в популяции начнётся так называемый дрейф генов, то есть случайное ненаправленное измерение частот аллелей с падением генетического разнообразия популяции. К дрейфу генов приводят и катастрофическое снижение численности особей в популяции вследствие пожаров, наводнений, засухи, сильных морозов или антропогенных воздействий, Умысл этого явления впервые истолковали С. Райт и Н.П. Дубинин, При утрате части генофонда вследствие дрейфа генов увеличивается генетическая однородность популяции, возрастает ее : омозиготность.Дрейф генов играет важную роль в микроэволюционных процессах. Если по некоторым причинам от большой популяции растений за счет дисперсии диаспор отделится небольшой ее фрагмент, особи которого будут несколько отличаться по набору генов от основной популяции, то на базе такого фрагмента сформируется новая популяция, генетически отличная от исходной. Это явление называют эффектом основателя. В другом случае основная часть популяции может быть полностью уничтоженной (пожары, лесные рубки и т.п.), но сохранится ее небольшая часть опять-таки с набором аллелей, несколько отличающимся от среднего типа. На базе этой сохранившейся части сформируется новая популяция, уже генетически неполностью идентичная исходной. Этот случай носит название эффекта бутылочного горлышка. На схеме рис. 10.1 представлены основные механизмы,
%
і
I
і
%
\
В
А
Б
Г
распределение аллелей в направление действие
исходных популяциях стрессового фактора
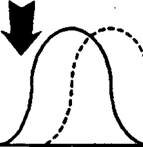
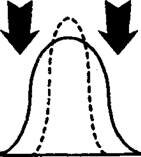
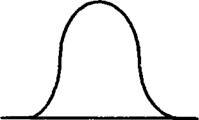
Рис.
фактора на популяцию:
А - исходная популяция;
Б - стресс подавляет один на крайних аллелей;
В - стресс подавляет крайние аллели;
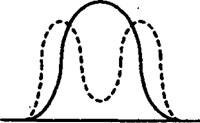
Г — стресс подавляет типичные особи с типичным набором аллелей и дает преимущества крайним формам
которые ведут к изменениям частот конкретных аллелей при раз ных воздействиях на популяцию.
В ходе микроэволюционного процесса в каждой популяции отбираются те генотипы, которые обеспечивают наилучшую адаптацию их носителей к местным условиям. В итоге микроэволю- ционный процесс ведет к появлению новых видов растений, но- зых жизненных форми новых стратегий растений, ориентированных на выживание и процветание.
Еще по теме Закон Харли - Вайнберга:
- Уравнение Харди—Вайнберга
- Биогенетический закон
- ЗАКОНЫ И ОБЪЯСНЕНИЯ
- Различные интерпретации «Лесного Закона»
- Открытие Г. Менделем законов наследования
- 1-15. Морфология и законы эволюции
- 4.6.1. Понятие об «эмпирических законах» и элементарной логической задаче
- Второй закон. Локализация события
- 13.2. СООТНОШЕНИЕ ОНТО- И ФИЛОГЕНЕЗА 13.2.1. Закон зародышевого сходства
- Для организма следует ожидать новых законов
- Физические законы основаны на атомной статистике и поэтому только приблизительны
- 1-7. Изменчивость, отбор и законы познания
- 5. Законы наследования. Дарвинизм по Уоллесу
- 3.5. Закон сохранения материи и энергии
- Основана ли жизнь на законах физики?
- Работа организма требует соблюдения точных физических законов
- Ф.МЮЛЛЕР -Э ГЕККЕЛЬ. ОСНОВНОЙ БИОГЕНЕТИЧЕСКИЙ ЗАКОН ИЗБРАННЫЕ РАБОТЫ, 1940