Регуляция каст со стороны семьи
У муравьев, ос и пчел на касты делятся только самки, а у термитов - и самки, и самцы. В этом разделе рассматривается способность репродуктивных (половых) особей регулировать образование особей той или иной касты.
Вообще такая регуляция может быть связана- с определенным периодом
в жизненном цикле или развитии семьи и
- с определенным временем года, подходящим для успешного выведения половых форм. В простых сообществах проблема касты решается доминированием маток над особями, равными ей по рангу; если же матка не способна сохранить свое положение вследствие какого-либо повреждения, возраста или снижения плодовитости, то это открывает дорогу ее преемницам. Например, у Mischocyttarus
drewseni- неотропического представителя Polistinae-по мере роста семьи стареющая матка уже не может заставлять всех появляющихся на свет самок становиться рабочими, так что появляются репродуктивные самки (и одновременно с ними самцы) (Jeanne, 1972). У другой осы, Metapolybia aztecoides, молодая самка скорее всего станет рабочей особью, если в момент ее появления на свет в семье уже имеются матки; в противном случае она будет копулировать и станет маткой (West-Eberhard, 1978). У одной из Halictinae-.Evy/amv calceatus - в первом потомстве бывает несколько самок, которые крупнее других и остаются в гнезде. Если самка-основательница терпит неудачу, они способны копулировать и сразу начать откладывать яйца (Platcaux-Quenu, 1978,
- . У других видов Evylaeus, например у Е. marginatus, в детерминировании касты участвуют самцы, так как без самцов самки не могут копулировать и у них не развивается обычное репродуктивное поведение, свойственное молодым маткам. Поэтому в таких случаях появление самцов приводит к развитию самок (Brian, 1980).
Существует и совсем другая проблема: в каком состоянии должна находиться семья, чтобы переключиться с производства рабочих особей на выведение репродуктивных самок? Таким состоянием может быть высокое отношение числа рабочих особей к числу личинок.
Это отношение начнет снижаться только тогда, когда плодовитость матки начнет падать или приблизится к своему пределу, поэтому оно не служит хорошим показателем состояния, наиболее подходящего для размножения. Тем не менее у многих примитивных общественных насекомых этот показатель, по-видимому, эффективен, например у видов Bombus, строящих восковые пакеты. На довольно ранних стадиях развития сообщества, если экспериментально повысить отношение рабочие/личинки, добавив рабочих особей или удалив часть личинок, появляются репродуктивные самки. В данном случае это всего лишь сверхкрупные самки, которые интерградируют по величине к рабочим особям, но у которых возникают репродуктивная диапауза и склонность к копуляции (Brian, 1980). Из данных Cumber (1949, цит. по Brian, 1965) следует, однако, что при появлении половых особей откладка яиц самкой не снижается. Кроме того, у B.pascuorum, который тоже строит восковые пакеты, численность колоний до образования половых особей варьирует в пределах от 30 до 150 особей, так что ввиду наличия у них одной матки отношение рабочие/матки вряд ли может быть решающим фактором. И в самом деле, экспериментальные исследования показали, что в этой группе видов Bombus матки не влияют на поведение рабочих особей, связанное с выращиванием расплода. Таким образом, можно, по-видимому, заключить, что главные факторы, от которых зависит формирование половых самок,- это запасы корма вне гнезда и отношение рабочие/личинки в гнезде.У ос Polistes умеренного пояса каста самок при надобности детерминируется уже после выведения имаго под влиянием таких сезонных факторов, как температура и длина дня (см. Brian,
- . Возможно, хотя еще твердо не установлено, что здесь играет роль также доминирование матки, так как в начале сезона размножения экспериментально повышенное отношение рабочие/личинки приводит к появлению только крупных рабочих особей, но не репродуктивных самок. У осы P. canadensis место данной особи в иерархии имеет большее значение, чем сезонные изменения, очень незначительные в тропиках, и с ростом семьи доля репродуктивных самок постепенно возрастает (West-Eberhard, 1969). С этой тенденцией коррелируют отношение рабочие/матки (P. canadensis - полигинный вид), отношение рабочие/личинки и, конечно, средний возраст маток. Таким образом, у более примитивных пчел и ос жизнеспособную популяцию рабочих особей создает та матка, которая обеспечивает их „занятость" путем непрерывных требований корма и регулярной высокой интенсивности откладки яиц.
У Lasioglossum zephyrum в группе осемененных сестринских особей в результате их взаимодействия через несколько дней выделяется матка (доминирующая самка). Она начинает
вести себя определенным образом: проявляет более выраженное стремление к дальнейшей копуляции, к образованию и откладке яиц, подталкивает рабочих особей, которые стараются избегать ее, сидит около входа в гнездо у конца очереди, образуемой здесь рабочими, а также занимает другие позиции, позволяющие ей контролировать перемещения рабочих и их деятельность. Она совершает также много коротких посещений ячеек - выясняет, в каком состоянии находится их строительство и снабжение кормом; полученные сведения позднее могут быть использованы для того, чтобы координировать и направлять деятельность рабочих особей (Buckle, 1982).
У шмелей (Bombus), которые все время кормят своих личинок отрыгиваемой пищевой смесью, матки, несомненно, регулируют кастовую принадлежность своих потомков женского пола.
Эволюция этого нового регулирующего фактора отодвигает на второй план роль отношения рабочие/личинки, поскольку меньшее количество корма просто означает меньшее число репродуктивных самок, а не большее число рабочих. Непрерывное наблюдение за личинками (Pendrell, Plowright, 1981) дает рабочим особям возможность распознавать, какие из них склонны развиваться в репродуктивных самок; затем рабочие путем непрерывного и обильного кормления стимулируют полное развитие потенциальных задатков таких личинок (в тех случаях, когда матка ослаблена или отсутствует) или же (если матка полна сил) противодействуют этому; они либо разбавляют корм, либо вовсе перестают кормить личинок, так что последние не могут полностью реализовать программу своего развития и потому становятся рабочими особями.
Какого поведения будут придерживаться рабочие по отношению к личинкам женского иола, зависит от частоты их контактов со своей маткой; по-видимому, им нужен подлинный физический контакт с ней по крайней мере раз в день (Roseler, 1977; Roseler, Roseler, 1977, 1978). Поэтому способность матки Bombus контролировать деятельность рабочих особей зависит от числа последних (строго говоря, от отношения рабочие/матка) и от активности матки и ее способности вырабатывать феромоны, которая с возрастом снижается и, вероятно, зависит от длины дня и температуры. Кроме того, возможно, что рабочие особи разного возраста и разной величины неодинаково восприимчивы к контактам с маткой. Главная особенность матки как реіулятора, видимо, состоит в том, что она способна суммировать все эти факторы и резко изменять поведение, с тем чтобы в определенное время года все усилия семьи полностью и бесповоротно переключались на выведение репродуктивных самок вместо рабочих особей.Meliponini не прибегают к системе непрерывного наблюдения и кормления, как высшие представители рода Bombus. Они просто устраивают для самок крупные ячейки с большими запасами корма, но у них должна быть какая-то не известная пока система, которая регулирует число этих ячеек. Молодым репродуктивным самкам, возможно, обеспечивается заботливое попечение в их ячейках, и их кормят через дырочки, выпуская наружу только тогда, когда рабочие особи „решают", что нужно устроить „конкурс" самок. В этом конкурсе, описанном Imperatriz-Fonseca (1977) у Paratrigona subnuda, рабочие особи вначале, по-видимому, бывают пристрастны к некоторым самкам, но позднее принимают его исход как должное. По всей вероятности, рабочие


Фото 28.
Передача корма от одних рабочих особей Vespula sp. другим на соте, ячейки которого содержат куколок и запечатаны шелком и картоном. Обратите внимание на личинок, расположенных по периферии.Фото 29. Рабочие особи медоносной пчель. образующие „свиту“ вокруг своей матки. Вс. они обращены к матке головами, и их усик: вытянуты вперед. [Фото 28 и 29-J. В. Free.
особи всегда чувствуют функциональную потенцию матки-быть может, по агрессивности, которую она проявляет во время церемонии откладки яиц, или по частоте этого события, или, возможно, благодаря вырабатываемым ею феромонам. У некоторых видов Melipona репродуктивные самки имеют такие же размеры, как рабочие особи (этих самок принято называть микрогинами). Как полагают, личинки, склонные к развитию в репродуктивных самок, генетически чем-то отличаются от других, но им необходимо несколько больше корма - только в этом случае доля таких самок может достигнуть максимума (25% всего потомства). Данные в пользу этого представления суммировал Brian (1980).
У ос Vespinae производство репродуктивных самок начинается тогда, когда впервые строятся большие ячейки. Стимулом для этого служит подача маткой нового сигнала, а не затухание прежнего сигнала. Хотя матка все время проявляет неоспоримое доминирование, активизирует рабочих особей и организует их жизнь в сообществе с помощью феромона контактного действия (Landolt et al., 1977), на определенном этапе своей жизни она начинает выделять какое-то новое вещество. Это противоположно тому, что происходит у Bombus, и, несомненно, входит в программу изменений, которую определяет в основном возраст, но на которую, безусловно, оказывают также влияние длина дня, наличие пищи и место в иерархии сообщества. У Vespa orientalis активным компонентом этого вещества служит лактон с 16 атомами углерода, возможно продукт мандибулярной железы. Было показано, что он стимулирует строительство ячеек, даже если его предъявляют рабочим в отсутствие матки на каком-либо инертном, неструктурированном материале, например на бумаге (Ishay, 1975).
У Apis mellifera наступление зрелости семьи знаменуется устройством маточников; постройка этих ячеек начинается в том случае, если весной плотность пчел высока (Lensky, Slabezky,
- . Самка обычно способна остановить строительство „запасных" ячеек в зоне расплода с помощью нескольких механизмов; самый важный из них-это, несомненно, секреция мандибулярными железами ненасыщенной жирной кислоты с низкой летучестью - 9-оксодекановой кислоты (9-ОДК). Это вещество распространяют в сообществе рабочие особи, которые образуют эллипс, или „свиту", вокруг матки всякий раз, когда она останавливается. Они прикасаются к ней своими усиками и ротовыми частями, а затем убегают в состоянии гиперактивности и передают полученный химический сигнал другим рабочим пчелам (Ferguson, Free, 1980; Seeley,
1979). У А. сегапа рабочие особи прикасаются к матке менее часто и обмен информацией между рабочими соответственно меньше. 9-ОДК сама по себе привлекательна для рабочих пчел, хотя они не образуют „свиту" вокруг кусочка смоченной этим веществом бумаги или пробки, как они это делают вокруг живой матки. Это вещество, несомненно, эффективно на протяжении большей части года, однако в мае и июне-главном сезоне размножения-необходимы очень большие искусственные дозы, чтобы предотвратить постройку маточников. В это время в дополнение к 9-ОДК могут потребоваться другие факторы, например следовой феромон, вырабатываемый тарзальной железой и оставляемый лапками матки на краях сотов.
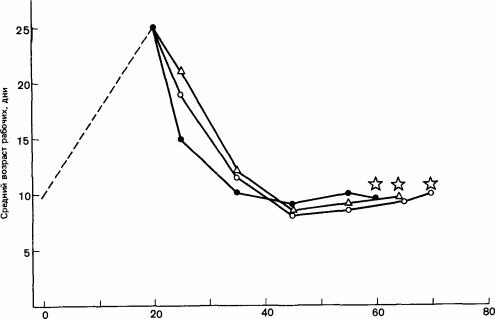
Время после роения, дни
Рис. 11.3. Цикл размножения африканской расы медоносной пчелы. Роение происходит через 20 - 30 дней (звездочки) после того, как средний возраст рабочих снижается до 10 дней. (Winston, 1972.)
Плодущие матки оставляют гораздо более обильный бесцветный маслянистый след, чем может оставить рабочая особь (Lensky, Slabezki, 1981). Сам по себе этот след не очень активно подавляет даже запечатывание маточников, но в сочетании с секретом мандибулярных желез он очень эффективен. В этом участвует также феромон, выделяемый брюшком, который, по-видимому, водорастворим, но о котором пока мало что известно (Brian, 1980). У A. mellifera, как и у А. сегапа, в роевом клубе информация о присутствии матки передается вверх, но не вниз по цепи пчел, сцепленных друг с другом ногами, причем передача сигнала, как бы она ни происходила, может преодолевать разрывы в 2, но не в 4 мм! Сигнал передавался и в экспериментах с раздавленными матками, так что он, очевидно, имеет химическую, а не поведенческую природу, но при этом не требуется непрерывной передачи химического вещества вверх по цепи рабочих пчел (Rajashekharappa, 1979). Даже молодые репродуктивные самки, еще находясь в своих ячейках, могут подавлять строительство других маточников (Boch, 1979).
Уинстон и Тэйлор (Winston, Taylor, 1980), специально изучавшие стимулы для выращивания репродуктивных самок у Apis mellifera (африканская раса, живущая в Южной Америке на 5° с. ш.), установили, что целый комплекс таких стимулов суммируется, достигая некоторого порогового триггерного уровня. Главную роль среди них играют численность семьи, площадь сотов и число взрослых особей; значения всех этих факторов должны быть высокими и должны возрастать, хотя пчелы могут не заполнять всю полость гнезда. По данным Уинстона и Тэйлора, в ульях объемом 20-25 л пчелы переставали строить маточники, когда они покрывали 0,8-1 м2 поверхности сотов. Необходимы были также большой процент молодых пчел, сильная их скученность в зоне, занятой расплодом, где пчелы стараются протиснуться внутрь, и локальный избыток пищи (рис. 11.3). Эти факторы приводят к разбавлению феромонов матки и к временному снижению или прекращению их секреции. На 5° с. ш., где длина дня почти не изменяется, а зимы не бывает вовсе, у пчел наблюдаются циклы продолжительностью около 50 дней. После того как ячейки построены и матка отложила в них яйца, она не может прекратить кормление личинок. Однако после окукливания и запечатывания ячеек она может непосредственно атаковать их, и если только она проделает в ячейке отверстие, рабочие особи разрушают ее до конца. В больших перенаселенных ульях рабочие пчелы особенно активно защищают ячейки с куколками от нападений матки и первых появляющихся репродуктивных самок; вслед за их появлением происходит многократное роение (Simpson, 1974). Рабочие пчелы исполняют своего рода вибрирующий танец, который, как считают, успокаивает этих агрессивных самок (Fletcher, 1978а). У муравьев самки тоже играют важную роль в регулировании размножения, влияя на образование как молодых самок, так и самцов. У Odontomachus haematoides (Ponerinae) рабочие особи при наличии в гнезде самки игнорируют требование корма со стороны крупных личинок, которые склонны к превращению в репродуктивных самок и из которых экспериментатор может вырастить таких самок. Иными словами, у муравьев в отличие от медоносной пчелы самка может регулировать кормление. Решающее значение, несомненно, имеет количество корма на поздних стадиях личиночного развития, а не его качество (Colombel, 1978, цит. но Brian, 1980). У Eciton удаление самки приводит к высокой смертности мелких личинок, из которых должны развиться рабочие особи; это, вероятно, объясняется плохим уходом за ними и созреванием нескольких новых самок, предположительно из обычных личинок женского пола, которым уделяется особое внимание и отдается переработанный корм. В засушливый сезон, к которому, несомненно, чувствительна вся семья, самку изолируют в той части находящегося в покое сообщества, где нет расплода, а в другой его части, иногда отделенной какой-нибудь физической преградой вроде бревна, выращивается обширное потомство из личинок мужского и нескольких личинок женского пола. Последние становятся репродуктивными самками. Границы между этими двумя зонами нередко вызывают конфликты между рабочими особями. Предполагается, что яйца, из которых разовьются самки, откладываются самкой незадолго до разделения на зоны; неизвестно, однако, откладываются ли ,,самцовые“ яйца самкой или рабочими особями, лишенными возможности контактировать с самкой (Schneirla, 1971). Аналогичный процесс имеет место у Апотта (Raignier, 1972). Создается впечатление, что всю эту систему создают рабочие в ответ на какое-то сезонное изменение, но первые небольшие сдвиги, возможно, происходят в самке и связаны с утратой ею способности эффективно вырабатывать какой-то феромон или откладывать яйца, а скорее всего-с тем и другим.
У представителей неспециализированного рода Myrmica семьи достигают репродуктивного состояния вследствие ослабления влияния матки на поведение рабочих-как прямого влияния, так и в результате образования рабочих особей, не реагирующих на ее присутствие.
Семьи М. ruginodis созревают при численности рабочих особей от 900 до 3000. Как полагают, рост их численности на этом не прекращается; он может продолжаться и после того, как появятся половые особи, подобно тому как это происходит у многих организмов. Поскольку М.ruginodis-полигинный вид, высокая вариабельность численности при достижении зрелости объясняется числом самок. Критическая цифра составляет примерно 1000 рабочих на одну самку (Brian, 1965, р. 57; Brian et al., 1981b). Первоначально предполагалось, что у М. rubra, еще более полигинного вида, самки собираются вместе и образуют единую группу; так оно бывает весной, когда они выходят наверх, чтобы погреться, но в общей форме это неверно: они агрегируют лишь в незначительной степени. В сущности, матки относятся друг к другу враждебно и рассеиваются по разным участкам гнезда, где за ними лучше ухаживают и где они в большей мере общаются с рабочими особями, чем если бы они, подобно маткам Apis, передвигались вместе со свитой (Evesham, 1982).
Основа этого „эффекта разбавления"-то, что самке необходимо время от времени прикоснуться к каждой рабочей особи, чтобы сохранить возможность управлять ее поведением, связанным с выращиванием расплода. Мертвая самка обладает таким же действием, но после экстрагирования из нее липидов оно совершенно исчезает. Нет никаких указаний на то, что информация о самке в семье может передаваться от одной рабочей особи другой, хотя полностью исключить такую возможность нельзя (Brian, 1973с); может быть, такая передача осуществляется путем соприкосновения ротовых частей, как у медоносных пчел, или с помощью какого-либо иного церемониального поведения, эффект которого не передается через тонкую металлическую сетку с такими мелкими ячейками, что рабочая особь пролезть через них не может.
У Myrmica, так же как и у Odontomachus, регуляция состоит в недостаточном кормлении личинок, из которых получаются репродуктивные самки,-по-видимому, их кормят водянистой отрыгиваемой пищей, а не слегка пережеванной животной пищей (табл. 11.1). У них наблюдаются также иные действия (не обнаруженные у Ponerinae): они упорно и энергично кусают брюшко личинки, что вызывает метаморфоз, приводящий к образованию рабочей особи. Эта агрессивная деятельность (ибо она производит именно такое впечатление) против репродуктивных самок возбуждается в рабочих особях легче и распространяется от самки сильнее-как во времени, так и в пространстве,-чем подавление активности, связанной с кормлением. Если рассматривать ее как результат передачи некоторой информации, то она проникает в сообщество глубже, чем информация, запрещающая уделять внимание личинкам репродуктивных самок. Последняя хорошо действует только вблизи от самки, но в смысле регуляции каст она более надежна, чем агрессивное поведение, которое сильно варьирует по своей интенсивности. Однако торможение кормления легче снимается в результате социальных взаимодействий, и с ростом сообщества этот используемый самкой регулирующий фактор исчезает в первую очередь. Самки, выступая в роли стимуляторов трудовой деятельности, заставляют также личинок женского пола пропускать одну важную стадию роста и
Таблица 11.1. Влияние маток у муравьев Myrmica на поведение рабочих особей по отношению к личинкам и откладке яиц
| Тип поведения | В присутствии самок | В отсутствие самок |
| (1) По отношению к личинкам, из которых развиваются рабочие особи | Кормилицы активно кормят этих личинок в ущерб другим; личинки метаморфизи- руют, превращаясь в рабочих особей | Этими личинками пренебрегают, и они перестают расти, достигнув середины третьего возраста |
| (2) По отношению к лабильным личинкам женского пола | Рабочие особи (главным образом фуражиры) нападают на этих личинок весной, когда они сигнализируют о своей способности превратиться в репродуктивных самок при помощи кутикуляр- ной секреции брюшка | И рабочие особи, и фуражиры кормят этих личинок твердой пищей, богатой белками, ухаживая одновременно за ограниченной их группой |
| (3) По отношению к личинкам мужского пола | Ими пренебрегают, но на них не нападают | Их кормят лучше, чем личинок, из которых должны развиться рабочие особи, но хуже, чем лабильных личинок женского пола, способных дать репродуктивных самок |
| (4) В отношении образования и откладки яиц | До тех пор, пока самка откладывает яйца или пока в гнезде есть кучка яиц, независимо от того, дают ли они самцов или самок, молодые рабочие-кормилицы откладывают неполноценные яйца, пригодные только в качестве корма (кормовые яйца). Очень молодые рабочие особи (в возрасте менее трех недель) откладывают репродуктивные яйца, которые формируются до того, как эти рабочие начинают реагировать на присутствие самки | Молодые рабочие особи откладывают крупные, полноценные репродуктивные яйца, которые не оплодотворены, но способны развиваться партено- генетически, давая самцов |
метаморфизировагь в рабочих особей. Эта проблема будет рассмотрена в главе 12.
Рабочие особи, испытавшие на себе воздействие маток вскоре после появления на свет, лучше реагируют на них в дальнейшем, когда начинают выкармливать личинок (Brian, Evesham, 1982; Carr, 1962); они обращаются с лабильными в смысле будущей кастовой принадлежности личинками более решительно: при наличии самки морят их голодом, а в ее отсутствие - перекармливают. Первые 3 или 4 недели, очевидно, имеют решающее значение для такого обучения рабочих; неясно, однако, надо ли рабочим особям просто познакомиться со своей самкой, или они нуждаются в ее помощи, чтобы научиться отрыгивать пищу личинкам, или же, что более сложно, общение с самкой как-то вводит их в общественный образ жизни. Smeeton (1981) показал, что они „рождаются" в физиологическом состоянии, соответствующем одиночному образу жизни, и будут откладывать репродуктивные яйца, если не испытают влияния плодущих самок. Поэтому если при недостатке в сообществе новых рабочих особей в нем существует дефицит самок (т. е. отношение рабочие/самки высокое), то весьма вероятно, что впоследствии многие рабочие не будут реагировать на влияние последних.
У Leptothorax nylanderi воздействия семьи также играют большую роль, чем снабжение пищей и отношение рабочие/личинки, причем самки варьируют по своей способности подавлять развитие новых самок; так же как и у Myrmica, их власть увеличивается с возрастом и достигает пика в начале каждого сезона (Plateaux, 1970, 1971). Наряду с различиями в силе воздействия самки в зависимости от времени года и возраста сами рабочие особи варьируют по своей реактивности, отчасти, несомненно, по причинам, указанным выше. Более крупные и более старые рабочие особи, главным образом фуражиры, сильнее реагируют на самок, чем мелкие и молодые, которые по большей части несут функции кормилиц. С ростом семьи средние размеры рабочих особей вначале возрастают. Это, в частности, хорошо установлено у Myrmica ruginodis (Brian, Brian, 1951), у M.sabuleti (Elmes, Wardlaw, 1982a) и у Lasius niger (Boomsma et al., 1982). У Lasius доля самок среди крылатых половых особей возрастает с увеличением размеров рабочих до предела, равного примерно 0,8, а затем возрастание прекращается, вероятно из-за того, что образование самок нарушает воспроизводство рабочих. Таким образом, по мере роста семьи самка получает все большую поддержку, так как крупные старые рабочие особи в присутствии самок нападают на личинок, а в их отсутствие возвращаются к работам в гнезде и помогают кормить личинок. Это может быть обусловлено сенсибилизацией рабочих особей к влиянию самки в начале их жизни в семье меньших размеров, где частота их контактов с самкой была выше. Однако по мере того, как семья растет со все большей скоростью, молодых рабочих особей становится все больше, и уже по одной этой причине они будут недостаточно сенсибилизированы, поскольку их контакты с самкой (или самками) не могут быть частыми. Такая структура характерна для семьи, постоянно увеличивающейся в геометрической прогрессии, а не для семьи с постоянной численностью, и это именно то состояние, в котором воспроизведение должно достигать максимума (Brian, Jones, 1980). Таким образом, созревание семьи обусловлено большим приростом рабочих особей,
недостаточно сенсибилизированных к влияниям, исходящим от их самки, в результате чего они не обеспечивают достаточного кормления личинок, потенциально способных дать репродуктивных самок.
У Myrmica и Leptothorax наблюдается еще одно изменение, на этот раз касающееся самок и связанное с их возрастом. Для личинок, вылупляющихся из яиц, которые самки откладывают в первый год, вероятность метаморфизировать в рабочих особей выше, чем для личинок из яиц более старых самок; эти „поздние" личинки чаще развиваются в направлении репродуктивных самок, хотя это развитие не обязательно доходит до конца. Имеющиеся в настоящее время данные не подтверждают мнения о существовании яиц двух типов (см. выше); скорее, все яйца укладываются в непрерывный ряд-от таких, в которые изначально заложен ранний метаморфоз, приводящий к формированию рабочих особей даже при низких температурах (20°С), до таких, которые метаморфизируют в рабочих особей только в том случае, если рабочие-кормилицы уделяют им активное внимание весной, в присутствии самок и при стимуляции со стороны последних. Без такого внимания они впадают в диапаузу, если только температура не окажется высокой (например, выше 25°С). Диапаузирующие личинки перезимовывают и медленно (при низкой температуре) приобретают способность развиваться в репродуктивных самок; но они лабильны в отношении касты, т. е. их развитие в самок не детерминировано. Самки по мере своего созревания откладывают все меньше яиц, детерминированных к развитию в рабочих особей, и больше яиц, лабильных в отношении касты, которые потенциально способны дать самок и только ожидают подходящих социальных условий, чтобы созреть. Однако даже у самок, которым больше года, весной есть короткий период, когда они могут откладывать некоторое количество яиц для выведения рабочих. Важную роль здесь, вероятно, играет быстрота откладки, а не малые размеры яиц; это позволяет думать, что наклонность к развитию в рабочих особей обусловлена каким-то дефектом, нарушающим эмбриогенез и ростовые потенции личинок, но не подавляющим развитие полностью, как в случае кормовых яиц. Индуцируемая самкой детерминация яиц в отношении касты неизвестна у пчел и ос, но обычна у муравьев; ясно, что это результат каких-то процессов, происходящих в организме самки, передаваемый ею сообществу через яйца (Brian, 1980).
У Monomorium pharaonis этот аспект развития очень четко выражен. Молодые самки откладывают яйца, большая часть которых дает мелких рабочих особей (хотя сами яйца крупные); созревая, самки начинают откладывать яйца, из которых могут развиваться крупные рабочие особи, и, наконец, к концу 120-дневного цикла -яйца, дающие личинок, предрасположенных к развитию в самок. Эти личинки выглядят по-иному: они толстые, лишены волосков и покрыты золотистыми точками. Матки откладывают некоторое количество неоплодотворенных яиц, и из них развиваются самцы. После производства половых особей и копуляции в гнезде цикл возобновляется (Petersen-Braun,
1977, цит. по Brian, 1980). Важно, однако, указать, что в этом цикле есть гарантии от ошибок, так как только крупные, произведенные в середине цикла рабочие особи вместе со стареющими самками будут выращивать личинок, дающих самок; в других случаях их убивают и поедают, возвращая их компоненты в общий круговорот веществ. Самки, помимо своего материнского воздействия на яйца, оказывают сильное ингибирующее влияние на производство половых особей через поведение рабочих; это влияние недостаточно тонко, чтобы переключить личинок с пути развития самок на путь рабочих особей, как у Myrmica,-оно просто приводит к их уничтожению (Edwards, 1982). Рабочие особи узнают своих самок по нелетучему липиду на поверхности их тела, как и у Myrmica (Bemdt, 1977; Bemdt,
Nitschmann, 1979; Brian, 1980); кроме того, самки выделяют из расположенной за заднем конце тела экзокринной железы неоцембрен, и это служит для рабочих особей сигналом начать нападение на личинок, предрасположенных к развитию в самок (Edwards, 1982). Таким образом, все черты социального развития, свойственные обитающим в умеренном поясе видам Myrmica, есть и у тропического Monomorium с циклическим развитием; у Myrmica бывает по два цикла в год-один с половыми особями, а другой без них, тогда как у Monomorium ежегодно бывает по три полных цикла, в каждом из которых производятся половые особи. Как рабочие особи или самки Monomorium узнают личинок, из которых должны развиться половые особи, не известно, хотя для человека они явно различаются как по форме тела, так и по текстуре поверхности. Кроме того, их поверхность, несомненно, обладает какими-то химическими особенностями, как и у муравья Myrmica rubra, кожные железы которого выделяют жир, плавящийся при 20-25°С (Brian, 1980). Разные виды Myrmica относятся нейтрально к лабильным личинкам не своего вида, но из личинок собственного вида выращивают только тех, которые должны дать рабочих особей. Можно предполагать, что у всех видов данного рода личинки, обладающие потенциями для развития в половых особей, имеют один и тот же опознавательный признак (Brian, 1975; Elmes, Wardlaw, 1982с). У Solenopsis invicta личинки, развивающиеся в половых особей, возможно, секретируют триолеин. Если пропитать этим липидом фильтровальную бумагу и разрезать ее на кусочки величиной с личинок, то рабочие особи будут собирать эти кусочки и складывать их вместе со своими предкуколками, дающими половых особей; они их не съедают, хотя муравьи этого вида очень любят жирную пищу (Bigley, Vinson,
1975).
Зрелые самки Pheidolepallidula-другого представителя Мугтісіпае-тоже откладывают более лабильные яйца, в основном ранней весной.
Вылупляющиеся из этих яиц личинки имеют характерную грушевидную форму, и их выкармливают только в том случае, если в гнезде имеется примерно 1400 рабочих особей, так как у этого вида никогда не бывает больше одной самки (Passera, 1980а). Вполне возможно, что материнское влияние на эти яйца обусловлено ювенильным гормоном, содержащимся в гемолимфе, так как обработанные этим гормоном самки откладывают более лабильные яйца (Passera, Suzzoni, 1979).
Виды Myrmica различаются по уровню, которого успевают достигнуть в своем развитии эти лабильные личинки до наступления зимы. У М. scabrinodis они неотличимы от других личинок по величине и внешнему виду, но у М. rubra и особенно у М. sulcinodis иногда бывают гораздо крупнее, чем зимующие личинки рабочих особей (Elmes, Wardlaw, 1981). Тем не менее они все еще способны сделать „шаг в сторону" и стать рабочими, в отличие от зимующих личинок Tetramorium caespitum, дающих самок: эти личинки настолько крупнее личинок рабочих особей, что в этом случае возможность переключения на другую касту сомнительна (см. Brian, 1974а). Может быть, этот эволюционно продвинутый представитель Myrmicinae, подобно Pheidole и Monomorium, просто уничтожает и поедает личинок, кастовая принадлежность которых детерминирована, а не „трансформирует" личинок, лабильных в отношении касты. Возможно, что причина утраты рабочими яичников-то, что путь их развития определяется уже на стадии яйца.
У Plagiolepispygmaea (Formicinae) регуляцию каст осуществляет самка; пока она имеет сперму в сперматеке и сохраняет активность, она может стимулировать у личинок рабочих особей ранний метаморфоз, как у только что рассмотренных Myrmicinae; всякая личинка, зашедшая в своем развитии слишком далеко, уничтожается и возвращается в круговорот веществ. По-видимому, стимулятором служит при этом какой-то феромон, так как если смочить самку ацетоном, то ее воздействие на личинок прекращается (Passera, 1974, 1980). Лабильные личинки зимуют, находясь в третьем возрасте (т. е. на более продвинутой стадии, чем личинки, дающие рабочих особей или самцов); поэтому можно думать, что они, вероятно, развиваются из яиц, отложенных в начале предыдущего лета, и что для полного развития им необходим период прохладной зимней погоды, как и у Myrmica. Таким образом, их жизненный цикл связан со сменой времен года (Brian, 1977).
Прежде чем обсуждать лесных муравьев, нам следует рассмотреть особый случай социальных паразитов. Plagiolepis grassei - паразит P. pygmaea - способен предотвращать развитие новых самок у своего другого хозяина (L. acervorum). sublaevis (Leptothoracini, Myrmicinae) могут заставить уже осемененных самок одного из своих хозяев (Leptothorax muscorum) прекратить откладку яиц, но не оказывают такого действия на самок своего другого хозяина (L. acervorum). Создается впечатление, что паразит регулирует образование половых особей у своего хозяина. У этого паразита матки часто бывают бескрылыми, хотя яичники и сперматеки у них вполне развиты; по данным Buschinger (1978), бескрылость обусловлена у них единичным доминантным геном. Возможно, что и самки Strongylognathus testaceus- паразита Tetramorium caespitum - подавляют образование половых самок у вида-хозяина. По-видимому, все эти паразиты продуцируют большее количество ингибирующего феромона, чем самка хозяина, или же их феромон действует сильнее.
Известно, что Formica polyctena откладывает яйца двух типов и что яиц промежуточных типов у них мало или нет вовсе (Gosswald, Bier, 1953; Bier,
1953). Яйца, откладываемые в начале каждого сезона, лабильны, но в природе из них развиваются только самки, так как плодущие самки, отложив эти яйца в теплых поверхностных камерах, уходят в глубь гнезда, предоставляя рабочим одним выращивать расплод. Это, конечно, тем легче, чем больше семья, и может рассматриваться как один из путей ослабления власти самки у данного вида. После периода отдыха в глубинной зоне гнезда самки откладывают мелкие яйца, из которых развиваются рабочие особи независимо от того, находятся ли эти яйца вблизи самки или нет; они детерминированы к развитию в рабочих особей. Однако Гёссвальд и Бир показали, что если их перенести в гнездо другого, менее полигинного вида F.pratensis, то из них могут развиться репродуктивные самки; таким образом, они „детерминированы" только в естественной обстановке-в гнезде собственного вида. Различие между яйцами этих двух типов обусловлено температурой и скоростью оогенеза; помимо прочего, мелкие яйца не содержат РНК'. В чем состоит влияние самок на выращивание рабочих особей, остается неизвестным, однако кажется вероятным, что самки заставляют рабочих убивать и возвращать в общий круговорот веществ личинок, способных развиться в самок, поскольку таких личинок можно легко распознать начиная со второго возраста: у них меньше волосков и более крупные головы (Schmidt, 1974).
Сходная регуляция существует и у термитов: если удалить репродуктивную пару, то формируются замещающие особи (псевдогины).
У низших термитов они образуются из личинок и нимф, у которых половые органы созревают столь преждевременно, что крылья и глаза не развиваются должным образом. У Termitidae это невозможно, и крылатые формы должны развиваться у них обычным путем в нормальные сроки, как и у муравьев. После этого они образуют пары, замещающие удаленных репродуктивных особей. Некоторые термиты, по-видимому, различаются по своей способности прибегать к одному или другому из этих способов. Если они находятся на стадии небольшой молодой семьи с множеством мелких личинок, то они создают замещающих особей, а на стадии зрелой семьи-крылатых взрослых особей. Здесь снова проявляется элемент сезонности: перезимовавшие личинки наиболее чувствительны и с наибольшей вероятностью превращаются в нимф, а затем, после дальнейших линек,- в имаго
1 Следует учитывать, что цитируемые работы были сделаны в 50-х годах, когда методика определения РНК была очень грубой -Прим. ред.
(Brian, 1979а). Так же как и у медоносной пчелы, образуется избыток замещающих особей; они вступают в драки, а рабочие приканчивают раненых, с тем чтобы сохранилась одна пара. Действительно ли при этом происходит борьба не на жизнь, а на смерть, приводящая к отбору наиболее приспособленных, проверить трудно.
В регуляции замещения участвуют оба пола, хотя их роли довольно различны. Казалось бы, каждый пол должен регулировать собственное замещение, однако это не так. У Calotermes удаление самца действительно ведет к образованию новых самцов, но при удалении самки образуются не только новые самки, но и новые самцы. Очевидно, самец не способен самостоятельно подавлять свое замещение; ему необходима помощь самки или, как это ни странно, помощь другого самца (Luscher, 1964). Наличие компаньона даже того же пола явно играет важную роль в функционировании пары: деятельность репродуктивной пары не сводится к созданию зигот. Вполне возможно, что половые особи более активно обмениваются информацией друг с другом, чем с представителями других каст, а это может означать более тесную связь и вытекающую отсюда более сильную стимуляцию метаболизма. У Neotermes jouteli опять-гаки разнополые пары могут регулировать замещение особей и того и другого пола, однако однополые пары, состоящие как из двух самок, так и из двух самцов, могут подавлять всех самок, но не полностью всех самцов. Этот процесс явно связан с сотрудничеством между взрослыми особями и зависит не от одних только ингибиторов, специфичных в отношении пола (Nagin, 1972). Конечно, высказывались мнения об участии в этом феромонов, распространяемых с пищей (Luscher, 1974, 1976), и о важной роли контакта (Springhetti, 1972). Если покрыть всего термита, кроме ротовых частей, лаком, то это не нарушает регуляторную функцию; в настоящее время полагают, что одна из желез головы вырабатывает ингибирующие феромоны, передаваемые через рот. Это, несомненно, слишком упрощенное представление: в такой регуляции участвует и какой-то поведенческий сигнал (Brian, 1979а).
Еще по теме Регуляция каст со стороны семьи:
- Морфогенез каст Термиты и муравьи
- ДВЕ СТОРОНЫ МЕДАЛИ
- Co стороны дедушки или бабушки вашим предком была обезьяна?
- ДВЕ СТОРОНЫ ОДНОЙ МЕДАЛИ: РОЛЬ АТМОСФЕРНЫХ АЭРОЗОЛЕЙ В ФОРМИРОВАНИИ КЛИМАТА
- Процессы регуляции в клетке
- 3.6.6.3. Регуляция экспрессии генов у прокариот
- Регуляция активности генов и белков
- 8.3.2. Эмбриональная регуляция
- Организация семьи
- Метаболическая регуляция
- Эпигенетическая регуляция онтогенеза
- ФАКТОРЫ, ВЛИЯЮЩИЕ НА ПРОДУКТИВНОСТЬ СЕМЬИ
- ГЛАВА 15. РОСТ ПЧЕЛИНОЙ СЕМЬИ
- СОСТАВ ПЧЕЛИНОЙ СЕМЬИ
- 6-8* Многослойная регуляция
- Основание семьи
- Регуляция генной активности
- Генетическая регуляция онтогенеза
- Рефлекторная регуляция положения тела
- 4. 4. Биологические механизмы регуляции численности