Самцы общественных перепончатокрылых
У термитов образование самцов - проблема, не связанная с поведением; подобно самкам, они формируются из зигот, и пол у них определяется хромосомами. Это не означает, конечно, что отношение полов должно быть равно единице, однако вклад обоих полов на стадии имаго неизбежно будет одинаковым.
Проблемы возникают лишь в том случае, когда этот механизм не обеспечивает оптимального численного отношения между стерильными кастами (рабочие/солдаты), как у Termitidae и в других семействах, где пол влияет на вероятность образования солдат вследствие генетических различий между самцами и самками.Для перепончатокрылых характерен гапло-диплоидный механизм определения пола, и большинство диплоидов-самки, при условии строгого аутбридинга. Гаплоиды-всегда самцы и развиваются из неоплодотворенных яиц; эти яйца могут быть отложены либо самками, либо рабочими особями. Поскольку у общественных перепончатокрылых, за исключением очень немногих видов муравьев, самцы никогда не выполняют никаких работ в гнезде и не защищают его, их производство представляет собой напрасное расходование энергии и других ресурсов и его необходимо контролировать, с тем чтобы самцы достигали зрелости только в тех семьях, которые могут выдержать такие затраты, или в семьях, которым может оказаться выгодным замедлить скорость своего роста. Это означает, что при некоторых обстоятельствах самцов можно рассматривать как фактор, замедляющий рост семьи на ранних этапах ее развития, а реализуемую ими отрицательную обратную связь - как стабилизирующее воздействие. Конечно, они во всякое время служат носителями генов, потенциально способными распространять гены самки или рабочей особи.
Примитивные пчелы, сообщества которых все еще представляют собой всего лишь ассоциации осемененных самок, часто производят смешанное в отношении пола потомство. Матка у них просто крупнее и старше других особей; она вмешивается в их поведение и стимулирует их активность по выкармливанию расплода, например у Lasioglossum zephyrus (Breed, Gamboa, 1977).
Нередко другие самки, образующие семью вместе с данной маткой, остаются неосемененными и могут откладывать только такие яйца, из которых развиваются самцы; хотя они не могут заменить матку, они могут нанести ущерб ее шансам на успех, откладывая яйца и производя самцов на слишком раннем этапе жизни семьи. У Evylaeus calceatus матка может предотвращать копуляцию самцов с другими самками; для этого достаточно ее присутствия, в активном вмешательстве с ее стороны нет надобности. Если матке не удается прекратить откладку яиц другими самками или рабочими особями, она разрушает и поедает их яйца (например, у Bombus). Более изощренный способ состоит в том, чтобы морить их потомство голодом на личиночнойстадии, в результате чего из таких личинок развиваются мелкие особи, похожие на рабочих (например, у Evylaeus malachurus).
Обитающие в умеренных областях осы Polistes проявляют по всем этим чертам конвергентное сходство с пчелами (West-Eberhard, 1969, 1973). У P.fuscatus образование самцов сопровождается высокой скоростью откладки яиц, и хотя это может означать, что свою долю здесь вносят и рабочие особи, возможно и то, что матки откладывают яйца слишком быстро и поэтому некоторые из яиц остаются неоплодотворенными: так обстоит дело у паразитических перепончатокрылых, а по мнению Фландерса (Flanders, 1962, 1969),- и у общественных Hymenoptera.
У Metapolybia-одной из Polybiinae, образующей большие полигинные семьи, размножающиеся роением, матки настроены агрессивно по отношению к только что появившимся репродуктивным самкам, яичники которых могут развиться до того, как они примут участие в копуляции (West-Eberhard, 1977). Докопуляционная откладка яиц, дающих самцов, встречается у паразитических перепончатокрылых, и экспериментальное вмешательство на этой стадии может привести к регрессии яичников (Flanders, 1969). Однако у Protopolybia pumila рабочие особи и откладывают яйца, и сами же поедают как собственные яйца, так и яйца, отложенные матками, хотя последние охраняют свои яйца в течение некоторого времени после их откладки.
Высокое отношение рабочие/матки приводит к увеличению доли потомков мужского пола, что позволяет предполагать количественную регуляцию откладки яиц рабочими особями со стороны матки (Naumann, 1970; West-Eberhard, 1977). Эти роящиеся Polybiini со многими репродуктивными самками, рабочими и постоянно присутствующими в семье самцами, по-видимому, адаптировались к условиям тропического леса, где они часто подвергаются нападению птиц и муравьев. Такая разнообразная структура семьи дает им возможность быстро переселяться и восстанавливать семью где-нибудь в другом месте (Jeanne, 1979).Ос і астся еще кое-что добавить относительно таких видов Bombus, как пчела-чесальщица и виды, строящие восковые пакеты. У В. hypnorum матка в начале сезона нападает на рабочих особей, откладывающих яйца, очевидно распознавая их по запаху или поведению; позднее ситуация становится, по-видимому, хаотичной, и она нападает на всех без разбора (Roseler, Roseler,
1977). Эта общая агрессивность связана с неспособностью оплодотворить свои яйца. У высших Bombus доминирование мало проявляется до конца сезона; теперь достаточно хорошо установлено, что мандибулярная железа вырабатывает феромон, подавляющий откладку яиц рабочими особями (Honk et al., 1980). Этот феромон снижает активность их corpora allata, причем действие его распространяется по сенсорным и центральным нервным путям (Roseler et al., 1981). Это подтверждает данные о том, что матка оказывает влияние на corpora allata подчиненных ей рабочих (Roseler, 1977). Honk et al. указывают, что в этом, возможно, участвуют также поведенческие эффекты, поскольку рабочие особи толпятся около матки, откладывают вместе с ней яйца и поедают их друг у друга. Среди них выделяются более активные особи, они подчиняют себе других и в свою очередь подавляют у них откладку яиц (хотя не могут заставить их прекратить выращивание новых самок). Это более старые рабочие особи, и они могут жалить матку до тех пор, пока она не погибнет. Такая совместная откладка яиц с их последующим поеданием наводит на мысль, что яйцекладущая самка способна привлекать к себе других особей со сходными тенденциями, которые могут затем поедать свежие яйца и в результате откладывать больше собственных яиц; это напоминает откладку кормовых яиц у муравьев и у пчел Meliponinae.
У многих (но не у всех) видов безжалых пчел рабочие особи в отсутствие самки откладывают репродуктивные яйца; сразу после откладки яиц рабочие особи запечатывают ячейки (Beig, 1977), а позже из них выходят самцы. У одного вида Scaptotrigona postica рабочие особи откладывают репродуктивные яйца даже в нормальных семьях, управляемых матками. Эти яйца крупнее тех, которые откладывает матка; рабочая пчела-узурпатор кладет свое яйцо поверх корма, после того как матка отложила свое, а затем запечатывает ячейку.
Самцы вылупляются спустя 65 ч, а не 67 ч, как самки; они крупнее и активнее и очень скоро продырявливают женское яйцо и поедают его и личинку. Самцы всегда выходят из ячеек, содержащих два яйца (Beig, 1972). Рабочие особи откладывают эти яйца в обычные ячейки, в которых количество корма меньше среднего, хотя причины этого непонятны, если только не окажется, что самцам для удовлетворительного развития нужно меньше пищи (Beig et al., 1982). Цитируемые авторы полагают, что рабочие особи, откладывающие яйца, конкурируют за эти ячейки, после того как матка удалилась, и что количество корма не имеет значения - важно, чтобы над кормом оставалось большее пространство; но почему?
Успех личинок мужского пола в конкуренции с личинками женского пола был продемонстрирован в опытах на Myrmica (Brian, 1969; Brian, Carr, 1960). Трудно представить себе, чтобы в применявшихся искусственных гнездах личинка мужского пола в буквальном смысле поедала личинку женского пола; по-видимому, рабочие особи как-то распознают личинок. В тех случаях, когда они откладывают мужские яйца, вылупляющиеся личинки - их сыновья, так что можно предполагать наличие у них некоторого предпочтения, обусловленного генетическими факторами.
У Vespinae матка обладает неоспоримым превосходством в молодых семьях; она полностью подавляет откладку яиц рабочими особями и стимулирует социальное поведение. Было высказано предположение, что, как и у Bombus terrestris, она вырабатывает феромон, регулирующий развитие яичников у рабочих особей, но доказать это пока не удалось (West-Eberhard, 1977). Если удалить из семьи матку, воцаряется анархия: рабочие особи дерутся между собой и пренебрегают уходом за личинками; затем они устанавливают собственную иерархию, и тогда откладка яиц становится прерогативой лишь немногих особей высшего ранга, из яиц которых развиваются самцы. Обычно стареющая матка начинает откладывать неоплодотворенные яйца, утрачивает свое доминирующее положение и подвергается атакам со стороны бунтующих рабочих особей, которые изгоняют ее и принимаются производить самцов (Montagner, 1966). Несомненно, рабочие особи могут отложить много яиц, но какую долю яиц они производят, не установлено (Kugler et al. 1976). Специальных ячеек для самцов не строится, и у некоторых видов, например у Vespula vulgaris, самцы развиваются в ячейках для рабочих особей, а у других, например у Dolichovespula sylvestris,- в больших ячейках. Быть может, рабочие особи, откладывающие яйца, менее разборчивы, чем матки?
У медоносной пчелы для выведения самцов необходима предварительная постройка специальных „трутневых" ячеек, которые в семьях, растущих в природных условиях, создаются на периферических участках строящихся сотов, непосредственно перед тем как вдоль краев строятся маточники. Это делается только в достаточно больших сообществах, но как только эти ячейки построены, матка методично откладывает в них неоплодотворенные яйца, по многу за раз, а не по одному. Этот процесс регулируется маткой, так как оплодотворение этих яиц происходит крайне редко. Любые ошибки, допущенные маткой, исправляют рабочие пчелы: они очень тонко разбираются в личинках, легко распознают личинок диплоидных трутней или репродуктивных самок, оказавшихся не в своих ячейках, и уничтожают их. Нет сомнений, что у медоносной пчелы яйца, дающие самцов, обычно откладывает матка; однако в отсутствие матки некоторые рабочие особи откладывают яйца, хотя и неупорядоченным образом-как в маленькие, так и в большие ячейки. У большинства рас яйца, отложенные рабочими пчелами, дают самцов, однако у одной южноафриканской расы, Apis т. capensis, они способны производить диплоидные яйца, из которых развиваются рабочие особи, и фактически половые особи имеются в избытке.
Матка полностью подавляет откладку яиц рабочими особями. У этой пчелы не бывает „полуразвитых" кормовых яиц. Как полагают, 9-оксодекановая кислота, выделяемая маткой, ингибирует функцию corpora allata; однако для того, чтобы рабочие особи прекратили откладку яиц, необходимы очень большие дозы 9-ОДК, и местное применение ювенильного гормона не стимулирует функцию их яичников, а превращает их из кормилиц в сборщиц, у которых яичники не функционируют. Эта необычная реакция, возможно, зависит от дозы гормона. Создается впечатление, что у Apis corpora allata или не вырабатывают гонадотропного гормона, как у большинства насекомых, или же необычайно чувствительны к концентрации ювенильного гормона в крови. Удаление corpora allata у матки не препятствует нормальному образованию яиц (Rutz et al., 1976, цит. по Brian, 1980). Чем же тогда регулируется оогенез? Строительству трутневых ячеек благоприятствуют большие размеры семьи, а присутствие матки тормозит его. Судя по имеющимся данным (Free, Williams, 1975; Free, 1977), рабочие особи в состоянии определять соотношение трутневых и рабочих ячеек; возможно, что эта их способность имеет хеморецепторную природу, однако никаких химических различий между разными ячейками пока не обнаружено. Обычно трутневые ячейки строятся раньше, чем маточники, но в экспериментальных условиях маточники, содержащие личинок, индуцируют постройку трутневых ячеек, и между устройством ячеек этих двух типов, видимо, существует какая-то связь. После того как трутневые ячейки готовы, самка, очевидно, не может удержаться от того, чтобы не отложить в них весной неоплодотворенные яйца, однако рабочие особи уничтожают эти яйца как несвоевременные. В конце лета матка не станет откладывать яйца в трутневые ячейки, хотя рабочие пчелы строят их. Таким образом, самцы появляются между этими двумя периодами, т. е. в нужное время!
В августе, при подготовке к зиме, самцов обрекают на голод и изгоняют; рабочие особи жалят их, если в улье имеется матка, но не в ее отсутствие. Итак, вся система регуляции зависит от величины семьи, времени года и существования функционирующей матки, которая препятствует не только образованию яиц у рабочих особей, но и постройке ячеек для самцов. Только в больших семьях с достаточным количеством корма рабочие пчелы могут преодолеть такую оппозицию со стороны матки.
Во всех подсемействах муравьев известны примеры откладки рабочими особями яиц, из которых развиваются самцы; однако во многих случаях остается неясным, происходит ли это регулярно и повсеместно или только в случае необходимости и при участии лишь нескольких особей. У тех видов, у которых факт откладки яиц молодыми рабочими особями хорошо установлен, рабочие, вероятно, производят многих, если не всех, самцов, однако это нуждается в проверке. Теоретически такое предположение кажется правдоподобным (Oster, Wilson, 1978). В частности, соответствующие данные о Myrmica rubra весьма убедительны (Brian, 1969; Pearson, 1928b; Smeeton,
1981).
Plateaux (1981) использовал рецессивный ген pallens, обусловливающий у Leptothorax hylanderi бледную окраску и хорошо проявляющийся у самцов, чтобы показать, что самцов производят рабочие особи. Очевидно, что у тех видов, у которых рабочие обладают лишь рудиментарными яичниками, все неоплодотворенные яйца, так же как и оплодотворенные, должна откладывать самка. У таких видов, вероятно, существует период, когда сперматека закрыта; у Dorylinae этот период падает на сухой сезон, а у Tetramorium caespitum-ш конец лета. У последнего вида в каждой семье существует определенное численное соотношение между самцами и самками; поэтому возможно, что самка откладывает отдельными партиями смесь оплодотворенных и неоплодотворенных яиц. Formica polyctena весной производит яйца, из которых развиваются самцы, но опять-таки неясно, откладывают ли их рабочие или бескрылые репродуктивные самки; уже давно известно, что рабочие особи могут откладывать такие яйца, но это, конечно, не означает, что так оно обычно и происходит (Schmidt, 1974,
- . Такие яйца дают самцов только весной, когда молодые отдохнувшие рабочие особи несут функции кормилиц. Способы, которыми самки муравьев заставляют рабочих особей прекращать откладку репродуктивных яиц в неподходящее время года, варьируют.
Рис. 11.4. Зависимость числа репродуктивных яиц, откладываемых рабочими особями Myrmica rubra, от числа дней в неделю, которые их самки проводят в гнезде. (Brian et al., 1981.)
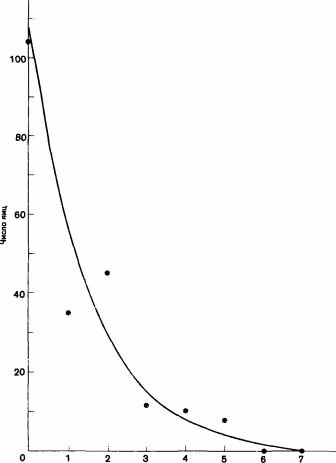
У Odontomachus haematodes рабочие могут оказывать сопротивление вмешательству самки; если их число достигает примерно 50, они образуют группу, которая не признает самку и изгоняет ее (Colombel, 1974). У других муравьев, например у Myrmica, самки сами откладывают много яиц. Это ведет к двум следствиям: во-первых, производя больше потомков женского пола, матки поддерживают „занятость" рабочих особей, и, во-вторых, они индуцируют откладку самими рабочими кормовых яиц. У Myrmica rubra откладка яиц маткой достигает пика ранней весной, еще до того как яичники перезимовавших рабочих особей будут способны вырабатывать желток; когда они начинают его вырабатывать, начинается вылунление личинок женского пола. Молодые рабочие, появившиеся в конце предыдущего лета, могут откладывать репродуктивные яйца даже в присутствии самок, которые вначале не влияют на них. Только достигнув 5-недельного возраста, если позволяет температура, они начинают откладывать кормовые яйца. Эти новые рабочие особи при 20‘ С могут начать откладку яиц в возрасте всего лишь трех недель; возможно, что эти яйца претерпевают некоторое развитие, когда рабочие особи еще находятся на стадии куколок (Smeeton, 1980, 1981). При полном отсутствии самок рабочие откладывают больше репродуктивных яиц при большей длине дня. При подходящей длине дня и температуре молодая самка может не только остановить производство рабочими репродуктивных яиц, но также использовать откладываемые вместо них кормовые яйца и продлить собственную откладку яиц. Для того чтобы осуществлять таким образом полный контроль, самка должна все время gt; дней в неделю, когда матка находится в гнезде
находиться в гнезде; даже в небольших гнездах каждый лишний день пребывания самки снижает наполовину число репродуктивных яиц, откладываемых рабочими особями (рис. 11.4) (Brian et al., 1981b). Иными словами, если самка проводит половину времени в одной небольшой камере гнезда, а половину-в другой, то только 10% яиц, отложенных рабочими особями, оказываются репродуктивными. Интересно отметить, что самка должна активно откладывать яйца и создать целый пакет яиц, чтобы
остановить производство самцов (Brian, Rigby, 1978), но производство половых самок может прекратиться при наличии даже мертвой самки! Подобным же образом недавно принятые самки могут оказаться неспособными остановить откладку рабочими особями некоторого количества репродуктивных яиц, однако они легко предотвращают образование новых самок (Winterbottom, 1981). Регуляция выведения самцов связана с каким-то более специфическим аспектом активности самки и, возможно, зависит от узнавания рабочими особями конкретных самок.
)iy концепцию подтверждает анализ модели Myrmica: как показала эта модель, в бедных ресурсами местообитаниях, способных обеспечить пищей только небольшие семьи, например у М. ruginodis на узких лесных полянах, производство самцов (в отличие от самок) не может быть подавлено. В таком состоянии может находиться любая семья, которая еще недостаточно хорошо обосновалась, например молодая; в первую очередь при этом будут выводиться самцы и лишь во вторую-самки, что слегка затормозит рост семьи. Кроме того, виды, у которых семьи делятся, будут производить потомство, состоящее из одних самцов, чтобы сбалансировать вклад обоих полов в следующее поколение (см. гл. 12).
Было высказано мнение, что это позволяет объяснить, каким образом у муравьев М. sabuleti, живущих в весьма однородном местообитании при сильной конкуренции со стороны других видов (Brian, 1972), могут быть семьи, производящие только самцов и имеющие в среднем примерно такие же размеры, как семьи, производящие и самцов, и самок. Plateax (1981) указывает, что семьи Leptothorax nylanderi, в которых не меньше 100 рабочих, производят только самцов.
До сих пор мы исходили из предположения, что рабочие особи регулируют производство самцов перед откладкой яиц, т. е. во время оогенеза. Это справедливо лишь отчасти, так как у Myrmica rubra личинки, развивающиеся в самцов, получают недостаточно корма, так же как и личинки, развивающиеся в самок, если плодущая самка находится где-то поблизости. Конечно, недостаточное кормление не может заставить их свернуть на путь развития в „рабочих“-самцов или привести к гибели; они просто развиваются в мелких половозрелых особей. Кроме того, личинки мужского пола не подвергаются таким нападениям, как личинки, из которых развиваются репродуктивные самки,-их не кусают, и они, по-видимому, не подают сигнала своим брюшком. Они преуспевают в конкуренции с личинками женского пола, как только что говорилось, и рабочие особи почти наверное отличают их и в отсутствие маток оказывают им предпочтение при кормлении (Brian, 1981).
У других видов самцов, несомненно, убивают на стадии личинок или даже яиц, если самка обладает сильным влиянием. Например, у Monomorium pharaonis личинок самцов можно обнаружить среди массы яиц на всех стадиях развития семьи, но ни одна из них не достигает зрелости (Peacock et al., 1954, цит. по Brian 1980). у Oecophylla рабочие особи откладывают очень крупные репродуктивные яйца, из которых развиваются самцы, и гораздо более мелкие яйца, которые тоже не оплодотворены и, развиваясь партеногенетически, дают самок; и те и другие откладываются в укромных уголках сложного гнезда, куда, по-видимому, не распространяется влияние самки (Ledoux, 1950). От чего зависит, будет ли партеногенез гаплоидным или диплоидным, не известно. У Cataglyphis cursor под влиянием самок яйца у рабочих особей рассасываются, но в отсутствие самок опять-таки имеет место партеногенез, как гаплоидный, так и диплоидный. Весной рабочие могут выращивать особей того и другого пола даже в присутствии самок из неоплодотворенных яиц; плодущие самки нужны только для того, чтобы активизировать рабочих особей и воздействовать на их поведение. Периоды партеногенетического размножения позволяют уменьшить расход ресурсов на самцов и распространить линию, которая на данный период оказалась хорошей в генетическом отношении; при этом периодически может происходить и половое размножение, приводящее к созданию новых признаков (Maynard Smith, 1978а).
Еще по теме Самцы общественных перепончатокрылых:
- ОБЩЕСТВЕННЫЕ НАСЕКОМЫЕ
- ОБЩЕСТВЕННЫЕ ОГОРОДЫ
- ГЛАВА 10. ПРОИСХОЖДЕНИЕ ОБЩЕСТВЕННОГО ОБРАЗА ЖИЗНИ У ПЧЕЛ
- РАЗДЕЛ III. ОБЩЕСТВЕННЫЙ ОБРАЗ ЖИЗНИ ПЧЕЛИНОЙ СЕМЬИ
- АЭРОИОНИФИКАЦИЯ ЖИЛЫХ И ОБЩЕСТВЕННЫХ ЗДАНИЙ И СООРУЖЕНИЙ V.I. ПСЕВДОАЭРОИОНЫ ВЫДЫХАЕМОГО ВОЗДУХА
- Брайен М.. Общественные насекомые: Экология и поведение: Пер. с англ.-М.: Мир,1986.-400с., ил., 1986
- Заключение
- Общие представления о созревании семьи
- Осы
- Теории индивидуального отбора
- Отбор родичей
- ШЕСТИНОГИЕ ИНТЕЛЛЕКТУАЛЫ