Лесные муравьи и термиты
Деревья открывают дополнительные возможности для экологической дифференциации видов, хотя они ослабляют или даже уничтожают наземный травяной покров, если их кроны смыкаются. Род Formica делится
. ... — sanguinea
І_ га/я
і r polyctena
L aquilonia/polyctem
aquilionia
¦— lugubris
[1— rufa/lugubris
1 pratensis
truncorum
exsecta
^ fusca
transcaucasica
cinerea
uratensis
родстве между собой. Среднее генетическое расстояние между популяциями одного вида равно 0,023, между видами-0,283, а между подродами-0,332. Все эти расстояния меньше, чем обычно бывает у животных (Pamilo et al., 1978, 1979). Группа F.rufa, в которую входит шесть видов (Kutter, 1977), отличается от других (см. дендрограмму на рис. 15.2), но эти виды либо генетически очень сходны, либо, дивергировав в разных географических
— rufibarbis
Рис. 15.2. Дендрограмма, показывающая родственные связи между евразийскими видами Formica (по данным о генетических расстояниях). (Pamilo et al., 1979.)
С
Рис. 15.3. Трансекта (с севера на юг) через долину реки Гуайдир, систематизированная так, чтобы показать местоположение гнезд лесных муравьев. Кружочки -Formica rufa; квадратики -F. lugubris. (Hughes, 1975.)
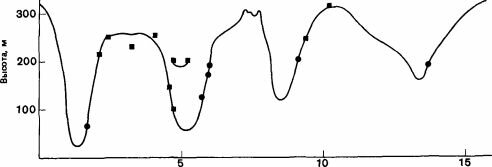
Расстояние, км
на несколько подродов, в число которых входят „рабовладельцы" Raptiformica, свободноживущие виды, которые строят маленькие холмики или не строят их вовсе и собирают корм в кустах (Serviformica), и рыжие лесные муравьи, строящие большие холмики, способные сохранять тепло, и фуражирующие на деревьях (Formica s. str.). Изучение 13 европейских видов показало, что они находятся в весьма тесном генетическом
областях, теперь все время скрещиваются между собой. Относительно генетических различий, необходимых для того, чтобы прекратить обмен генами и инициировать видообразование, практически ничего не известно. Гибридизация возможна, так как половые особи в Европе часто появляются в одно и то же время, и копуляцию между представителями разных видов наблюдали неоднократно.
F. truncorum и F.pratensis сильно отличаются от остальных, и их можно различать по морфологическим признакам (Длусский, 1967). У других четырех видов дискретных диагностических признаков найдено не было, и сходство между конспецифичными популяциями лишь ненамного больше сходства между видами. Любопытно отметить, что вид F. aquilonia в Шотландии, ныне представляющий собой изолированный фрагмент популяции, больше сходен с F. lugubris, обитающим чуть южнее, чем с финской популяцией F. aquilonia (отличающейся от шотландской формы по двум локусам). В свою очередь финская форма F. aquilonia более сходна с F. lugubris из южной части Финляндии, чем с шотландской F. aquilonia. Судя по генетическим данным, эти два вида, вероятно, дивергировали независимо один от другого на противоположных концах Европы[XII].
На Британских островах вид Formica lugubris в общем распространен севернее, a F. rufa южнее. Их ареалы соприкасаются и заходят один в другой в центральной части Уэльса, в Озерном округе, на йоркширских верещатниках и, возможно, на Пеннинских горах, но в Ирландии существует только F. lugubris (Breen, 1977, 1979). В Уэльсе вид F.rufa проявляет свою потребность в тепле тем, что он распространен от уровня моря до высоты 200 м на солнечных, обращенных на юг склонах (в большинстве случаев это . места заброшенных плавильных печей) с постоянными открытыми участками, окруженными дубовым мелколесьем (рис. 15.3). В тех местах, куда вторгается лес и кроны деревьев смыкаются, этот вид исчезает.
В тропическом дождевом лесу (Папуа-Новая Гвинея) обитают вместе два вида Leptomyrmex (Dolichoderinae). Крупные рабочие особи L. lugubris поедают разнообразных насекомых (в основном других муравьев), за которыми они охотятся с хорошо выраженных, хотя и временных дорог, а также занимаются сбором растительных материалов. Вид L.fragilis с более мелкими рабочими и семьями меньшей величины прокладывает много слабо выраженных троп, на которых муравьи собирают мелкие пищевые объекты, главным образом крылатых насекомых. Для L. lugubris помимо более крупных размеров и приверженности к питанию растительным материалом и муравьями характерны также охраняемые территории, на которые он не допускает рабочих из других семей своего вида или L.fragilis. Его гнезда расположены вблизи определенных деревьев, выделяющих эксудат, которые здесь немногочисленны. L. lugubris может занимать гнезда L.fragilis (обычно расположенные в центре территории с нечеткими границами) без всякой борьбы, буквально „выставляя" из них хозяев (Plowman, 1981). Ключевым ресурсом для этих муравьев служит, вероятно, надлежащий вид деревьев, дающих эксудат; любое из таких деревьев может быть захвачено муравьями L. lugubris, но при этом остается достаточно пространства и пищи и для L.fragilis.
Способу использования тропического дождевого леса на западе Малайзии двумя видами Macrotermes, составляющими вместе больше половины биомассы термитов в этой области, посвящена часть исследования Matsumoto и Abe (1979). У обоих видов есть грибные сады, которые они выращивают на кусочках растительного материала, главным образом сброшенных листьев;
однако в то время как М. carbonatus (0,33-0,43 г/м2) подбирается к ним сверху и строит гнездовые холмики, M.malaccensis (1,06 г/м2) подходит к ним снизу и строит гнезда под землей. Муравьи М. carbonatus предпочитают свежеопавшие листья, к которым они приходят ночью целыми колоннами, охраняемыми солдатами, нередко удаляясь по поверхности земли на 10 м от гнезда. М. malaccensis предпочитает прелые листья, хотя их может оказаться мало, но питается главным образом древесиной поваленных деревьев, а опад собирает только вблизи таких деревьев, построив над ним земляной навес и подходя к нему снизу. Таким образом, оба вида используют один и тот же пищевой ресурс - опавшую листву, однако они выбирают листья, находящиеся на разных стадиях разложения, и собирают их по-разному: один вид сверху и ночью, а другой-снизу и под защитой навеса. Активная популяция М. carbonatus может лишить М. malaccensis пищи в виде опавших листьев, однако у последнего вида еще останутся при этом поваленные деревья.
Источник: Брайен М.. Общественные насекомые: Экология и поведение: Пер. с англ.-М.: Мир,1986.-400с., ил.. 1986
Еще по теме Лесные муравьи и термиты:
- Расы муравьев и термитов
- Муравьи и термиты в грасслендах
- Морфогенез каст Термиты и муравьи
- ЭТИ HE БРЕЗГЛИВЫЕ ТЕРМИТЫ
- Термиты как разрушители растительных остатков
- Прибалочные лесные полосы
- ЛЕСНЫЕ ТИХОХОДЫ
- Муравьи
- Стокорегулнруюнще лесные полосы
- Тропический дождевой лес Осы, пчелы и термиты
- Муравьи пустынь
- Муравьи-листорезы
- Приовражные лесные полосы
- Тропические лесные красноземы
- МУРАВЬИ
- Луговые и лесные сообщества умеренной зоны Грассленды Европы
- Муравьи
- РОД ЛЕСНЫЕ АНТИЛОПЫ GENUS TRAGELAPHUS
- Муравьи в лесу
- Разведение муравьев — особый процесс